ГЛАВА 3. Географическая изменчивость антропогенетических данных
Впервые наследственность групп крови была выявлена в 1901–1904 годах, вскоре после того, как Де Фриз, Коренс и Чермак подтвердили генетические законы Менделя. В этой связи возник научный интерес к проблеме распределения новых антропологических факторов между этносами. В 1930 году Ф. Бернштайн предложил модель трехаллельной наследственности, согласно которой две доминантные аллели (р, q) по отдельности обусловливают, соответственно, вторую и третью группы крови, а вместе — четвертую. Рецессивный ген «r» в гомозиготном состоянии формирует первую группу крови, а в гетерозиготном — вторую и третью.
К середине 1960-х годов большинство этносов всех континентов, кроме Восточной Европы и Северо-Западной Азии были исследованы на предмет распределения доминантных, кодоминантных и рецессивных генов многих систем организма. Появилась новая возможность: в дополнение к традиционным характеристикам классической антропологии значительно расширить их количество за счет популяционно и генетически значимых. Если же выделить доминантную и рецессивную наследственность отдельных признаков полигенных систем, то при изучении популяционных и межэтнических различий можно использовать методы статистической генетики. В частности, мы широко использовали цвет глаз, оволосение средних фаланг пальцев рук, форму ушных раковин, отдельные признаки дерматоглифики и другие.
Подобная унификация широкого спектра антропогенетических и морфофизиологических систем человеческого организма позволяет подсчитать эмпирическое и теоретически ожидаемое распределение фенотипов и их аллелей по формуле Бернштайна.
Ряд археологических и палеоантропологических сообщений свидетельствуют о достаточно давнем заселении нашей территории человеком (в эпоху позднего палеолита) и о преемственности его родословной. Изученный нами генофонд и геногеография подтверждают местные корни беларуского этноса и позволяют считать его коренным на европейском пространстве. Так, размах частот генов системы АВ0 среди европейцев равняется 0,45–0,75 для г(О), 0,20–0,40 для р(А) и 0,05–0,20 для q(B) — при средних региональных значениях 0,580 ± 0,009; 0,236 ± 0,011; 0,184 ± 0,010. У беларусов эти показатели таковы: 0,56–0,64 (0,615 ± 0,009); 0,21–0,29 (0,241 ± 0,004); 0,12–0,17 (0,144 ± 0,007).
Согласно нашим данным, распределение их мало отличается от соседних этносов в пределах восточноевропейской зоны.
Для русских они составляют: 0,580 ± 0,006; 0,256 ± 0,006; 0,164 + 0,004.
Для украинцев: 0,590 ± 0,004; 0,261 ± 0,005; 0,149 + 0,003.
Для летувисов: 0,617 ± 0,007; 0,243 ± 0,011; 0,139 ± 0,020.
Для латышей: 0,573 ± 0,005; 0,260 + 0,007; 0,167 ± 0,006.
Для поляков: 0,560 ± 0,024; 0,280 ± 0,021; 0,160 ± 0,017.
Более высокие концентрации рецессивного гена г(О) как индикатора эндогамии выявлены у летувисов и беларусов; гена р(А) как гена европеоидности — у поляков, латышей и украинцев; q(B) как гена монголоидности — у русских и латышей. Таким образом, замеченные тенденции антропогенетической изменчивости свидетельствуют, что за фенотипическим подобием скрывается исторически обусловленная изменчивость.
Согласно данным о концентрации генов М и N, народы Европы разделяются натри большие географические зоны: западную, центральную и восточную. Беларуский этнос занимает промежуточное положение между центральной и восточной зонами.
В свою очередь, локальные варианты позволили выделить на территории Беларуси две основные зоны: юго-западную с частотой гена М более 60 % и северо-восточную с частотой менее 60 %. В популяциях Западного Полесья статистически достоверно чаще встречаются гомозиготы, что указывает на их относительную изоляцию в прошлом.
В Центральной и Восточной Европе концентрация гена М нарастает от средней региональной для Европейской историко-этнографической провинции (0,586 ± 0,012) к показателям (0,594 ± 0,012) — беларусы, (0,600 ± 0,024) — поляки, (0,606 ± 0,007) — украинцы, а у латышей — 0,672.
В конце 1940-х годов Р. Рейс и А. Винер (Race R., 1944; Wienier А., 1952) открыли новую изосерологическую систему «резус». Это существенно обогатило антропологическую генетику. Данная система достаточно сложно организована, ее детерминируют три пары аллельных генов, которые контролируют наследственность 8 гаплотипов. К настоящему времени точно установлено, что у жителей европейских стран наиболее часто встречаются следующие комбинации гаплотипов: CDe (50–56 %), CDE и cDE (10–15 %), сde (12–16 %). Последние же четыре гаплотипа либо редкие: cDe (2–3 %), cdE (1–2 %), либо очень редкие — CdE (менее 0,05 %).
Одни исследователи считают современный полиморфизм по резус-аллелям результатом мутаций и естественного отбора в пользу более жизнеспособных зигот, другие отдают преимущество дифференциации доисторических прапопуляций с их последующим смешением. Например, У. Бойд выделил палеоевропеоидную расу по наличию среди ее представителей до 50 % резус-отрицательных индивидов (Boyd W. C., 1950).
Между прочим, максимальные частоты рецессивного гена «d» отмечены в популяциях поляков (0,440), беларусов (0,396 ± 0,005), латышей (0,388 ± 0,10), летувисов (0, 381 ± 0,10) и украинцев (0,368 ± 0,011), наиболее удаленных от региональной средней Европейской провинции (0,304 ± 0,018).
По данным нашего исследования, концентрации гаплотипов резус у коренных беларусов имеют свое распределение, отличное от современного европейского. Так, среди нас преобладают гаплотипы «сde» и «CDe» с частотами 0,392 ± 0,035 и 0,324 ± 0,026, а гаплотипы «cdE» и «CDE» присутствуют в минимальном количестве (0,005 + 0,005 и 0,003 ± 0,006). Гаплотип «CdE» вообще не обнаружен. Именно такие генетические особенности позволяют сделать вывод об автохтонности и эпохальном инбридинге беларуских сельских популяций.
Межэтническая география имеет следующий вид. Если брать наиболее часто встречаемые гаплотипы, то частота «CDe» от минимальной концентрации у беларусов (0,3140) постепенно вырастает до значений 0,4189 (русские), 0,428 (латыши) и 0,435 (украинцы). Гаплотип рецессивных генов «сde» меньше у латышей и русских (0,342 и 0,362), но увеличивается в юго-западном направлении к беларусам (0,387) и украинцам (0,396). Резкой максимальностью редких гаплотипов «cDe» и «Cde» среди этносов-соседей выделяются именно беларусы — 0,122 и 0,039 по сравнению с латышами (0,027 и 0,027) и украинцами (0,035 и 0,007). Украинский этнос отличается минимумом гаплотипа cDE (0,095) и максимумом гаплотипа CDE (0,027) на всем пространстве Европейской историко-этнографической провинции.
Отметим также, что среди резус-отрицательных индивидов беларуско-украинского Полесья нами выявлены три самых редких в Европе гаплотипа. Они могли сохраниться еще со времен палеоевропеоидной расы (согласно классификации У. Бойда). Исследуя же жителей Минска (т. е. мегаполиса), мы при всей их потенциальной гетерогенности и гетерозиготности вообще не выявили 4 гаплотипа из 9 возможных. Этот факт наглядно показывает влияние естественного отбора если не в постнатальном, то в пренатальном[85] периоде онтогенеза.[86]
Мы изучали еще две системы — АВ0(Н) и Lewis(Leab), генетически тесно связанные между собой. Их антигены присутствуют почти во всех жидкостях организма. Первую мы обнаружили в слюне, а вторую — в эритроцитах. К сожалению, в этом случае можно провести сравнение только с русскими, потому что остальные наши соседи по этим системам вообще не изучались. Если по первому фактору беларусы и русские различаются мало (0,544 и 0,597), то по второму — намного больше (0,514 ± 0,017 и 0,381 ± 0,067). В сравнении со средним региональным показателем (0,437 ± 0,043) они занимают полярные позиции.
По иммунноглобулиновой системе Gm в границах Европейской историко-этнографической провинции исследованы 8 этносов, в том числе 3 восточнославянских. Распределение этнических частот ее гаплотипов выглядит следующим образом: русские — 0,177; беларусы — 0,162; украинцы — 0,144. Все три показателя ниже среднего регионального (0,254 ± 0,037).
Из систем антигенного полиморфизма, детерминированного двухаллельными локусами, изучались системы Р, Kell(Kk), Duffy(Fy). Гены первой локализованы на 22-й хромосоме, однако роль антигенов в физиологическом процессе до сего времени не установлена. Рецессивный ген «р», по нашей гипотезе, имеет свои особенности в географии распределения. На территории Европы пока что обследованы только 9 этносов в 65 локальных популяциях (из них 43 беларуские, 7 эстонских, 3 русских, 1 украинская, 1 польская). Средней региональной (0,549 ± 0,028) соответствует частота у русских (0,504 ± 0,063) в пределах средней ошибки. Для украинцев и эстонцев этот показатель одинаков (0,585 ± 0,025) и тоже близок к средней региональной величине. Польский и беларуский этносы выделяются: первый — минимальной концентрацией гена «р» (0,460), второй — максимальной (0,693 ± 0,032).
Антигены системы Kell характеризуются высокой иммунногенностью и занимают второе место после антигенов системы резус. Они совсем не зависят от других изосерологических групп крови, однако передаются по наследству согласно тем же генетическим законам, что и другие изоантигенные групповые признаки человека. Пока известно одно их существенное отличие — антигены «К» и «к» встречаются исключительно у европейских народов. Среди соседних этносов в направлении к региональной средней величине по рецессивному гену «к» приближаются, с одной стороны, русские (0,964 ± 0,004) и эстонцы (0,959 + 0,008), а с другой — украинцы (0,971 ± 0,003). Беларуси снова имеют свою специфику, хотя в данном случае и небольшую — 0,939 ± 0,020.
В начале выполнения всемирной программы «Геном человека» на 1-й хромосоме были открыты локусы Fya и Fyb, которые находятся в кодоминантных отношениях между собой. Антропогенетики используют систему Fya для определения генетического расстояния между представителями европеоидной и негроидной рас. Концентрация гена Fya у беларусов (0,494) практически не отличается от средней региональной для Европейской историко-географической провинции (0,488 ± 0,038). Между прочим, у эстонского этноса этот показатель минимальный (0,318 + 0,085) в пределах всей Северной Евразии.
***
Переходим к анализу систем генетико-биохимического полиморфизма, модели наследственности которых к настоящему времени хорошо разработаны. Это позволяет использовать их маркеры в популяционно-генетических исследованиях наряду с иммунологическими и серологическими системами. Наиболее широко в этом аспекте нами изучена система гаптоглобина — Нр. На распределение ее маркеров должны влиять и действительно влияют популяционные и этнические особенности. Теперь все антропологи пользуются формулой двухаллельной наследственности. Согласно ей, два аутосомных[87] кодоминантных аллеля Нр1 и Нр2 находятся на 16-й хромосоме и контролируют три фенотипных варианта: две гомозиготы (Нр1–1, Нр2–2) и гетерозиготу (Нр2–1).
Региональная средняя по гену Нр1 на пространстве Европейской историко-этнографической провинции (0,312 ± 0,011) почти не отличается от северной европейской средней величины (0,326 ± 0,009). Все соседние этносы вместе с беларуским по шкале межэтнического распределения частот генов Нр имеют повышенные концентрации. Ближе других к средней находятся латыши, даже не выходя за границы статистической ошибки (0,317). Далее идут летувисы (0,364 ± 0,012), русские (0,364 ± 0,17) и поляки (0,370), затем беларусы (0,409 ± 0,014) и украинцы (0,469 ± 0,008). Иначе говоря, при всей неоднородности распределения факторов гептоглобина на конкретной территории наблюдается общая тенденция скорее их меридиональной, чем широтной изменчивости.
Еще один сыворотковый белок из бета-глобулиновой фракции — трансферин (Тf, локус которого находится на 17-й хромосоме, — изучен среди 8 этносов Европы. Половина из них (беларусы, украинцы, русские, эстонцы) выступают довольно тесной группой.
По генам TfC, TfB, TfD почти идентичны беларусы (0,994; 0,004; 0,002) и украинцы (0,992; 0,002; 0,003), русские (0,988; 0,007; 0,005) и эстонцы (0,982; 0,008; 0,005). Но все они, будучи географически близкими друг другу, статистически достоверно не отличаются и от средней региональной (0,982 ± 0,004; 0,002 + 0,001; 0,015 ± 0,004). Сравнительно невысокий полиморфизм этой системы может свидетельствовать о том, что конкретный электрофорезный метод не в состоянии выявлять новые аллели.
Среди иммунных комплексов изучался еще один глобулин сыворотки крови — комплемент СЗ. Систему его фено- и генотипной изменчивости формируют два основных альтернативных гена (F и S) и несколько редких. Последние на территории Европейской историко-этнографической провинции пока что выявлены только у коми-пермяков (0,006) и русских (0,013). По концентрации же генов C3F и C3S именно беларуский этнос (0,131) находится в пределах среднего регионального показателя (0,141 ± 0,013). Вто же время русский этнос со своим минимальным показателем (0,105) ближе к североевропейской средней величине (0,087 ± 0,013). Наиболее отличается по концентрации гена C3F эстонский этнос (0,184), к которому только приближаются коми-пермяки (0,169) и хакасы (0,171). Несмотря на бимодальное распределение гена C3F в популяциях и этнических группах Европейской историко-географической провинции, их максимальное количество попадает в интервал с минимальными частотами генов. Полученные результаты не противоречат современному представлению о генетическом равновесии популяций, изученных с помощью статистического анализа Харди-Вайнберга.
На территории Беларуси мы, с помощью В. А. Спицына, исследовали одну популяцию коренных беларусов по системе Рр — сыворотковая щелочная фосфотаза. Возможны два генетически обусловленных варианта ее фенотипов — Рр1 и Рр2. Однако полностью генетическая структура этого маркера пока что не выяснена. Хромосомная локализация локуса Рр тоже еще не установлена. Число популяций, а тем более этносов, изученных на предмет фенотипного разнообразия остается небольшим, а в границах Европейской историко-этнографической провинции это только беларусы и русские. По частоте фенотипов Рр1 русские (0,797 ± 0,117) находятся намного дальше от североевропейской средней (0,581 ± 0,037), чем беларусы (0,630). Поданным В. А. Спицына и его коллег, модельные классовые интервалы популяционных и этнических распределений всех фенотипов Рр совпадают (Спицын В. А. и др., 2000). Выявленные довольно высокие значения ошибок в действительности могут свидетельствовать не только о значительной внутриэтнической дифференциации, но и о возможной неточности типологии.
***
Далее рассмотрим системы физиолого-генетического полиморфизма: зрительно-цветовую и вкусовую чувствительности. Возможность контроля функций цветового и вкусового анализаторов позволила расширить наши знания о генетических различиях людей в их восприятии окружающего мира.
Ныне существуют безукоризненные методы изучения структуры генофонда по единой схеме для всех генетических маркеров. Так, цветовая слепота определялась с помощью таблиц Рабкина, где она классифицируется как монохромозия, дихромозия, аномальная трихромозия. Последний синдром считается переходным к нормальной трихромозии у здоровых людей. Невосприятие красного цвета (протаномалия), зеленого (дейтераномалия) и синего (тританомалия) по отдельности встречается преимущественно среди мужчин и не всегда диагностируется. Дихроматы вообще не ощущают указанных выше цветов по отдельности и называются, соответственно, протанопами, дейтеранопами и тританопами. Монохроматы слепы на всю цветовую гамму полностью.
Генетико-популяционные исследования физиологического полиморфизма проведены на тех же индивидах, что и вся остальная антропологическая программа. Протаномалия и дейтераномалия контролируются двумя тесно сцепленными локусами на Х-хромосоме (Маккьюсик В. А., 1976). Детерминированный ими рецессивный признак дает возможность подсчитать частоту гена цветовой слепоты по соответствующей формуле. Еще в XIX веке была отмечена максимальная концентрация этих аномалий в прибалтийских губерниях России — до 9 % (Люблинский А. В., 1885). Согласно нашим данным, русские, украинцы и поляки вошли в группу с большей численностью слепых на зеленый цвет по сравнению со слепыми на красный. Рядом с ними оказались латыши, эстонцы и финны.
Из 29 обследованных популяций 17 беларуских изучены нами. Беларуский этнос (сельские популяции) показал самую высокую концентрацию генорецессивного «qan» — 0,140 ± 0,040. Интересная деталь: исследованные нами русские, живущие в Беларуси, по этому фактору (0,080) идентичны 9-тысячной выборке коренных русских (0,085 ± 0,006) в пределах статистической ошибки. Латыши, летувисы, украинцы, поляки тоже находятся на шкале высоких частот по отношению к средней региональной (0,073 ± 0,010). На карте Европейской историко-этнографической провинции с запада на восток, по мере усиления выраженности монголоидных черт, частота гена «qan» снижается. На территории Беларуси вектор его изменчивости имеет широтное направление.
В практике антропогенетических исследований последних десятилетий приобрел ценность еще один сенсорный маркер — вкусовая чувствительность к фенилтиокарбамиду (РТС). Соответствующие рецепторы ощущения горького вкуса РТС размещены на задней (дорсальной) поверхности языка. Вкусовой анализатор — эволюционно один из древнейших. Он сыграл значительную роль в филогенезе и естественном отборе в качестве «центра питания» (Тамар Г., 1976). В начале 30-х годов XX века полиморфизм данного признака был выявлен у людей и человекообразных обезьян. С этого началось популяционное и межэтническое изучение его распространения.
Постепенно с помощью индивидуальных и семейных исследований был установлен факт генетической наследственности вкуса к РТС. Система контролирует доминантную (Т) или рецессивную (t) передачу соответствующих генов от родителей детям. К концу 40-х годов была унифицирована методика обследования и определения минимальной (пороговой) концентрации РТС, при которой человек ощущает горький вкус. В результате было выявлено бимодальное распределение обследованных лиц на чувствительность к РТС. Наследственный фактор в ощущении других горьких веществ пока не обнаружен. Корреляция между вкусовым, зрительным и слуховым анализаторами тоже отсутствует.
Рассматриваемый признак тесно связан с расовыми особенностями, что позволяет широко пользоваться им в этногенетических исследованиях. Так, монголоидные популяции обладают чрезвычайно высокой чувствительностью, австралоидные же довольно низкой. На Евразийском континенте частота рецессивного гена «t» уменьшается в направлении с запада на восток. Его региональная средняя на пространстве Европейской историко-географической провинции равняется 0,590.
Среди соседних этносов вектор изменчивости сохраняется — минимум у поляков (0,530), максимум у русских (0,658 ± 0,30). Летувисы (0,565), беларусы (0,594 + 0,009) и украинцы (0,595) по вкусовой чувствительности почти не различаются.
***
Переходим к научным результатам изучения генофонда беларуского и соседнего населения согласно данным молекулярной гибридизации ДНК. Как следует из всего вышеизложенного, генетически детерминированные маркеры позволяют ответить на многие вопросы генетической географии популяций, взаимосвязей между этносами, их адаптационной зависимости. Но теперь появилась методологическая возможность на порядок расширить число совершенно новых маркеров за счет хромосомной ДНК.
При этом использование современных методов изучения ДНК-маркеров позволяет снять факторы влияния микроэволюции на генофонд. На фоне незначительной дифференциации в историческое время существования вида Homo sapiens естественный отбор не должен был затронуть изучаемые фракции ДНК.
В результате новейших научных разработок значительно увеличилась возможность использования молекулярно-генетических методов не только в биологической, но и в исторической (этнической) антропологии. Это новое научное направление предложено называть этногеномикой (Лимборская С. А., Микулич А. И. и др.). Генофонд народов восточноевропейского региона остается пока наименее изученным с этих позиций. Но материалов по отдельным популяциям Беларуси и беларускому этносу в целом уже достаточно для того, чтобы сравнивать их с генофондами других этносов. С этой целью использован полиморфизм гаплотипов ДНК в популяциях из разных географических регионов мира. Наибольшее разнообразие гаплотипов и дивергенция равновесия обнаружены у народов Центральной и Восточной Африки.
Как раз по исследованным гаплотипам локусов IGHJ, SCA, DRPLA, DM (соответственно 14, 6, 12 и 19-я хромосомы) выявлен чрезвычайно высокий уровень их гетерозиготности, что позволило уточнить древнюю историю трех восточнославянских этносов (с учетом эффективных размеров и степени разделенности локальных и региональных популяций коренного населения).
Данные по маркерам Y-хромосомы еще больше расширяют возможности молекулярной антропогенетики в проверке этноисторических гипотез. Наибольшую информационную значимость в плане исследования дифференциации людских популяций имеют те части генома, которые несут максимально нейтральную генетическую информацию. Например, величина индекса фиксации Fst по генетическому разнообразию находится в границах 0,0034–0,0057, что свидетельствует об этническом своеобразии региональных популяций (см. карту 2).
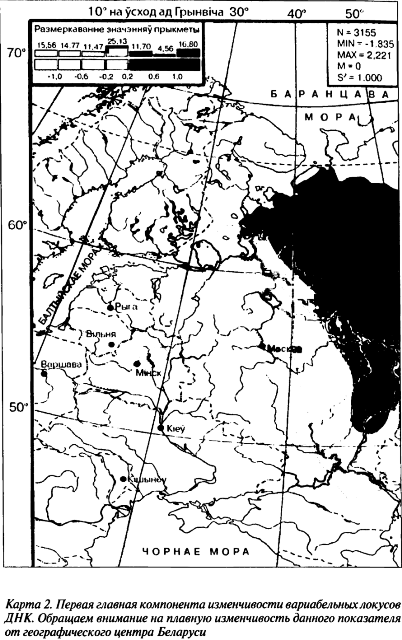
При изучении эволюции этносов дополнительно могут быть использованы особенности популяционного развития женской и мужской частей. В таком случае изучаются те маркеры ДНК, которые имеют однородную наследственность. К такому типу ДНК-маркеров принадлежат аллели Y-хромосомы. Они находятся только в геноме мужчины и передаются по мужской линии от отца к сыну. Дело в том, что часть Y-хромосомы, будучи уникальной, не способна к рекомбинации и передается через поколения в неизменном виде.
Совместно с украинскими и российскими коллегами мы исследовали пять микросателлитных маркеров. Считается, что именно они отражают исторические процессы. Медианные сетки гаплотипов позволяют проследить этническую взаимосвязь. Гаплотип 1 является мажорным[88] для новгородских русских (0,22), гаплотип 2 — для украинцев Киева (0,19). Пинские беларусы отличаются мажорной концентрацией обоих — 0,13 и 0,11. Эти результаты дают основание полагать, что беларуские популяции по указанным характеристикам более других соответствуют сообществу предков восточных славян.
Таким образом, изучение генофонда населения Беларуси в системе генофондов народов Европы позволяет определить не только наше геногеографическое положение, но и этногенетические связи с соседями. С этой целью мы используем обобщенный анализ всех характеристик, рассмотренных выше.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК