ГЛАВА 8. Этногеномика популяций Восточной Европы по ДНК-маркерам
До недавнего времени, изучая этническую геногеографию, мы пользовались преимущественно многочисленными выборками индивидуальных геномов. Диктовалось это сравнительно малым числом исследованных локусов. Нынешний молекулярно-биологический подход к изучению генетико-популяционной дифференциации позволяет значительно расширить количество геномных групп. Он существенно увеличивает достоверность полученных результатов по причине более эффективного нивелирования влияния возможных факторов микроэволюции. Последние имеют различную значимость как для отдельных генов, так и для отдельных популяций. Селекция человека как социального организма в историческое время генетически могла оставаться (и, скорее всего, оставалась) генетически нейтральной.
Все эти обстоятельства фактически свидетельствуют об одинаковой роли обобщенных генов при дифференциации генофондов по совокупности классических факторов.
«Исследование генофонда, то есть совокупности всех генов в популяции, является собственно популяционной задачей, при разрешении которой проблема функции гена второстепенна. Поэтому на завершающем этапе исследования не только возможно, но и необходимо получить обобщенные популяционно-генетические характеристики генов в системе генофонда».[107]
Именно с такими характеристиками мы и будем иметь дело в разделе этногеномики. Имеются в виду уровни генетической межпопуляционной дифференциации (Dst) и гетерозиготности популяций (Hs). Гетерозиготное различие между регионами как раз и отражает историко-культурные условия прошлого и демографические особенности населения соответствующих территорий.
В своих предыдущих публикациях мы широко пользовались также статистической категорией обобщенного гена — так называемым генетическим расстоянием между генофондами — d (квадратичное), которое, между прочим, весьма близко к среднему Fst (Edwards A. W., Cavalli-Sforza L. L., 1972). Связь действительно вполне очевидная. Она точно фиксирует усиление генетического разнообразия между популяциями соответствующим увеличением генетического расстояния между ними. Эти параметры достаточно объективны и линейно пропорциональны времени их дивергенции от популяций предков.
Однако точность этого соответствия будет наиболее высокой при том условии, что эффективно-репродукционные размеры популяций (N) остаются если не стабильными, то хотя бы без значительного (свыше четверти) притока иммигрантов (М). (Шереметьева В. А., 1975).
Уже предварительный анализ полиморфизма аллелей и гаплотипов позволил получить статистические результаты, отследить геногеографию обобщенных характеристик. Все это, вместе взятое, способствует дальнейшему поиску путей сохранения генодемографического разнообразия и возможной адаптивности.
В рамках отдельного беларуско-российского проекта ФФД № Г99Р-02 выполнен предварительный анализ по локусам DM, SCA1, IGHI, D1S80. Всего обследовано 8 выборок. Во всех популяциях суммарно выявлены практически все генотипы, характерные для европеоидной расы генотипа локуса 1GHI. Однако наиболее редкие из них встречаются в Иванове (10/11, 10/14, 11/12), Пинске (10/17, 7/10, 16/17), в Молодечно помимо двух последних еще и 8/17, в Мяделе 12/15, Климовичах 11/12. Из всех аллелей 14-й имеется только в Иванове, 7-й в Молодечно и Пинске, 15-й в Полоцке и Городке. 13-й аллель отсутствует во всей Центральной и Восточной Европе. Кроме того, у этнических беларусов отсутствует 11-й аллель. Пять аллелей (8-й, 9-й, 10-й, 12-й, 16-й) выявлены во всех изученных выборках.
На примере предыдущих материалов по изосерологической и морфофункциональной изменчивости нами установлен факт достаточно устойчивой генетической памяти в локальных сельских популяциях. По количеству 10-го и 12-го иммунноглобулиновых аллелей существует так называемое бимодальное распределение с модальной их частотой 0,71 + 0,03, в своеобразном виде — в границах экологических макровыборок: от 0,678 ± 0,03 на юге до 0,748 ± 0,01 на севере. Так, наибольшее число аллелей отсутствует в Придвинье — 7-й, 11-й, 14-й и 17-й; в Центральной Беларуси — 11-й, 14-й и 15-й, на Полесье — только два (11-й и 15-й).
Максимальная палитра изменчивости выявлена опять в Полесской геоморфологической провинции. Это явление — реликтовые особенности данной провинции — соотносится с нашими предыдущими открытиями редких гаплотипов в резус-системе. Матрица и дендрограмма локальных популяций сохраняют ту же меридиональную тенденцию. Видимо, в этом случае можно считать, что наряду с устойчивой генетической памятью существует и экологическая зависимость прапопуляций в процесе эволюции. Не исключено также миграционное влияние.
Таким образом, положено начало формированию соответствующей базы данных других аллелей, в частности, ответственных за иммуноглобулиновые факторы. Пока по этому локусу обследованы популяции трех сельских районов Минской области и трех средних городов (Молодечно, Полоцка, Пинска) в трех экологических провинциях Беларуси: Центральной, Придвинской и Полесской. Каждая из изученных популяций имеет свой характер распределения частот встречаемости аллелей IGHJ (HVR-Ig) (см. таблицу 1).

В настоящее время по данным литературы известно до восьми аллелей в разных этническо-расовых популяциях мира. Среди беларуских популяций идентифицированы все 8, но каждый из них имеет свои особенности. Так, в молодечненской и пинской популяциях не выявлен 15-й аллель, в полоцкой — 7-й и 17-й. Даже на этапе предварительного статистического анализа выявляется своя география распределения аллелей, по которой северные беларусы отличаются от южных и центральных. Наиболее часто встречаются аллели 10 и 12, что полностью совпадает с материалами литературных источников о русском и западноевропейском этносах. Это, пожалуй, то общее, что роднит европеоидов.
Особенности всегда выделяются по редким аллелям и генотипам. Например, генотипы 9–10, 9–12, 9–16 и 16–16 (4 из 19) не обнаружены в молодечненской популяции, а 8–9, 8–17 и 12–17 (3 из 19) отсутствуют в пинской популяции. Последняя в сравнении с остальными дополнительно выделяется почти вдвое более высокой концентрацией аллеля 16 (0,174). По наиболее редким гомозиготам 8–8 и 16–16 пинчуки тоже демонстрируют свое отличие. Если вспомнить выявленные нами раньше реликтовые гаплотипы резус-отрицательности — эпохи палеоевропеоидной расы в Беларуском Полесье — то подтверждается тезис об автохтонности полешуков с времен позднего неолита. Тем более, что степень гетерозиготности среди них значительно меньше даже по сравнению с населением центральной провинции (0,69 и 0,76).
Как видим, анализ даже одного высокополиморфного микросателлитного локуса предоставляет интересные данные о взаимозависимости между разными территориальными группами беларусов и позволяет проследить возможные миграционные потоки. Для получения более полной информации необходимо увеличение количества анализируемых локусов до 3–4. При высокой информативности они заменяют большинство классических маркерных систем (таких, как эритроцитные группы крови и белки кровяной плазмы). Только в таком случае ускоряется выполнение работ по анализу геногеографии в соответствии с экологическим районированием и современной экологически опасной ситуацией. С другой стороны, генотипоскопия на уровне индивида позволяет включить последнего в группу риска того или иного заболевания.
Одновременный анализ популяционных данных по обобщенным показателям поможет определить степени родства моноэтнических группировок (в границах Республики Беларусь), а также с популяциями смежных территорий. В этом смысле, как нам кажется, стоит еще раз обратить внимание на выполненное нами совместно с московскими коллегами сравнение по аллелям и генотипам локусов СА1 и CCR5 беларусов с рязанскими русскими и башкирами (Сломинский П. А. и др., 1997; Микулич А. И. и др., 1999). Из 14 аллелей локуса СА1 одинаково по 3 отсутствуют в популяциях и Беларуси, и Башкирии, причем два из них (30-й и 31-й аллели) не выявлены у обоих этносов. Кроме того, у беларусов отсутствует 32-й аллель, а у башкиров — 25-й. По частотам аллелей и генотипов гена CCR5 оказывается, что беларусы и башкиры наиболее отдалены друг от друга. Русские занимают в этом геногеографическом пространстве промежуточное положение (см. табл. 2).
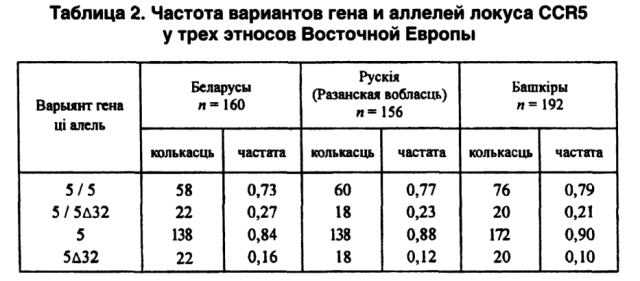
Таким образом, изучение каждой новой молекулярно-генетической системы при геногеографическом подходе к анализу популяционных данных (микроэволюционных процессов и истории формирования регионального населения) расширяет возможности современной антропогенетики. Именно геногеографическая интерпретация позволила В. В. Бунаку построить интерполяционные карты АВ0-распределения Центрально-Восточной Европы (Бунак В. В., 1969). Ю. Г. Рычков создал антрополого-генетическую школу (В. А. Шереметьева, В. В. Жукова, Е. В. Балановская, А. И. Микулич и другие), усилиями которой было начато издание многотомного фундаментального труда «Генофонд и геногеография народонаселения».[108]
Приведенные в первых двух томах гипотезы и суждения нашли свое подтверждение еще по одной из исследованных нами микросателлитных систем ДНК — локусе D1S80. Мы снова обращаем внимание на геногеографические особенности распределения полученных характеристик. Например, по наличию самых редких генотипов и аллелей значительно выделяется западнополесская популяция, где их выявлено соответственно 16 и 4, тогда как в центральнобеларуской — 8 и 2, в придвинской — 4 и 0. Как видим, и эта система не противоречит нашей концепции антропогенетической изменчивости беларуского населения в направлении с юга на север или наоборот.
Подсчеты для всей макропопуляции беларуского этноса показали наличие бимодальности распределения аллелей преимущественно за счет 24-го и 18-го (0,39 ± 0,03 и 0,26 + 0,03), что дает 2/3 всего массива данных. Из 21 выявленного аллеля еще только 4 имеют 5–7-процентную величину (31-й, 22-й, 28-й и 25-й). Встречаемость каждого из остальных 15 менее одного процента (табл. 3).
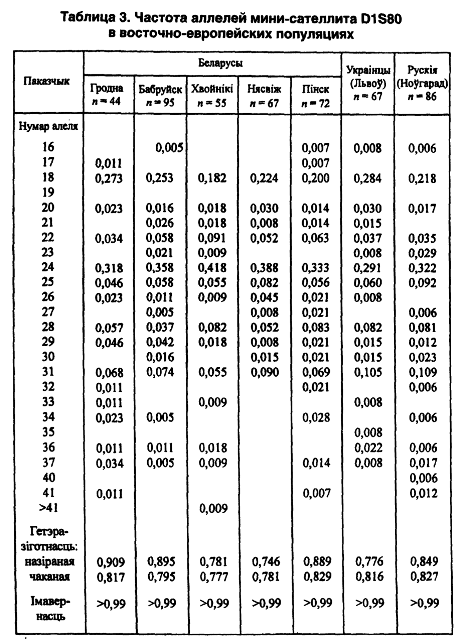
Однако на карте распространения 5-го аллеля локуса DM (карта 10) в восточнославянских популяциях выделяются 4 геногеографических региона с вектором северо-запад — юго-восток (Лимборская С. А. и др., 2002). Согласно данным указанной публикации, наблюдаемая и ожидаемая величины гетерозиготности беларуских популяций весьма близки друг другу, без отклонения от равновесия Харди-Вайнберга. И последнее замечание по данному локусу. Самые редкие аллели (17-й и 33-й) выявлены только у сельских (до 4-го колена и далее) беларусов и коренных немцев — у 0,7 и 0,5 процента соответственно. Все это косвенно свидетельствует о возможном воздействии стабилизационного отбора.
С целью решения задач этнической антропологии мы исследовали полиморфизм еще двух маркеров ДНК — SCA1 и CCR5. Из восточноевропейских этносов наряду с беларусами были обследованы башкиры и удмурты в первом случае, русские и башкиры — во втором. Изучались гетерозиготность и концентрации генотипов и аллелей. Наш этнос отличается от двух уральских максимальными частотами (10 % и более) гетерозиготных генотипов 22/27, 19/22, 19/28, а также 22/24, и гомозиготы 22/22 (в пределах 7–5 %). Гомозигота 19/19 у нас вообще не выявлена, тогда как среди представителей монголоидной расы она достигает 4 процентов. Для 13 аллелей локуса СА1 теоретически должен существовать 91 вариант генотипов. Но если среди волго-уральских популяций выявлена почти половина их, то в Беларуси — только четверть.
Популяции всех трех изучавшихся этносов по наиболее распространенным аллелям (19-й, 22-й, 27-й) статистически значительно различаются между собой. Показатель общей гетерозиготности наиболее возрастает от европеоидов (0,72) в сторону негроидов (0,86) (Лимборская С. А. и др., 2002).
Согласно результатам исследования по анализу генофонда маркера CCR5 среди беларуских, русских и башкирских популяций, его информационная значимость тоже достаточно высокая. Во всех изученных выборках распределение генотипов полностью соответствует закону Харди-Вайнберга (р<0,05). Концентрации всех генотипов и аллелей у беларусов и башкиров полярные, а у русских — промежуточные между ними. Здесь межэтнический вектор изменчивости имеет, согласно С. Лимборской, широтное направление. Аллельные варианты этого маркера по своим концентрациям существенно различаются в популяциях разных рас. Происходит это преимущественно за счет новых мутаций среди северных европейцев с последующим распространением на юг. Однако не исключена также связь с иммунитетом или с адаптационными возможностями как отдельных организмов, так и популяций в целом (Лимборская С. А. и др., 2002).

Более основательно исследована популяционно-генетическая особенность микросателлитного локуса DM гена миотанинпротеинкиназы (хромосома 19), являющегося причиной такого неврологического заболевания, как миотоническая дистрофия. На первый взгляд, нет существенных различий в распределении аллельных вариантов у разных территориальных групп беларусов по анализировавшимся выборкам (Попова С. Н., Микулич А. И. и др., 1999). Но только на первый взгляд. Локус DM в обычных условиях ведет себя нейтрально и поэтому становится пригодным для изучения популяций. Количество аллельных вариантов в разных популяциях беларусов неодинаково: от 9 до 12 из возможных 17.Что существенно — у беларусов выделены 4 аллельных повтора, пока еще не открытых в Европе: 19-й, 25-й, 27-й, 29-й. Однако такое явление не привело к отклонению равновесия Харди-Вайнберга (р< 0,89–0,99) и нет существенного различия (не более 2 %) между теоретической и наблюдаемой гетерозиготностью.
Выявленное бимодальное распределение повторов с пиками 5-го и 11–13-го аллелей фактически одинаковое как в беларуской популяции, так и в общеевропейской. Однако наличие у беларусов четырех перечисленных редких аллелей дополнительно свидетельствует об их древней автохтонности. К такому выводу мы пришли на основе подсчетов генетических расстояний между выборками с помощью пакета программ PopGene на базе алгоритма, предложенного М. Неем в 1978 году.
На соответствующей дендрограмме гродненская и хойникская популяции занимают полярные позиции, а Несвижская и бобруйская формируют промежуточный кластер. Почти меридиональный вектор изменчивости снова сохраняет свое географическое направление. Картографическая модель распределения в географическом пространстве 5-го аллеля как супермажорного (41 + 3) полностью подтверждает наш предыдущий аналитический прогноз 2002 года. Популяционные характеристики соседних этносов плавно перетекают от минимума у финскоязычных этносов к максимуму у московских русских и молдаван.
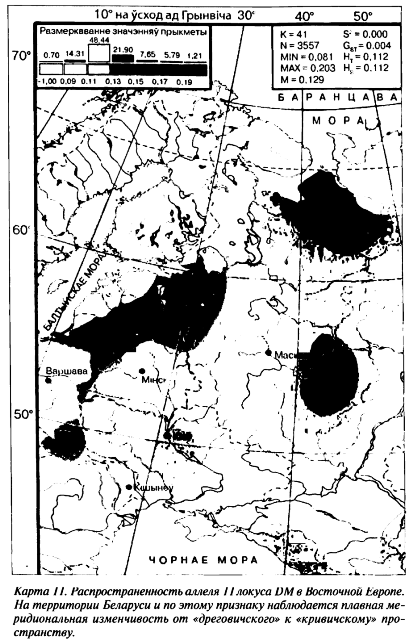
На геногеографической карте аллеля 11 локуса DM его минор[109] находится в центральной части Беларуского Полесья, а смежные концентрации объединяют южных беларусов с жителями Украинского Полесья и восточных беларусов с населением смежных регионов Российской Федерации (карта 11). Аллель 13 обладает противоположной частотной изменчивостью.
Согласно данным палеоантропологии, предки с этих территорий имели наибольшее число европеоидных признаков (Алексеев В. П., 1969). Археологи, в свою очередь, свидетельствуют о распространении здесь древностей зарубинецкой культуры (Третьяков П. Н., 1966; Седов В. В., 1982). Остается, правда, открытым вопрос: чьими предками могли быть носители зарубинецкой культуры — славян или балтов? Наш антропологический и популяционно-генетический материал свидетельствует в пользу тех и других.
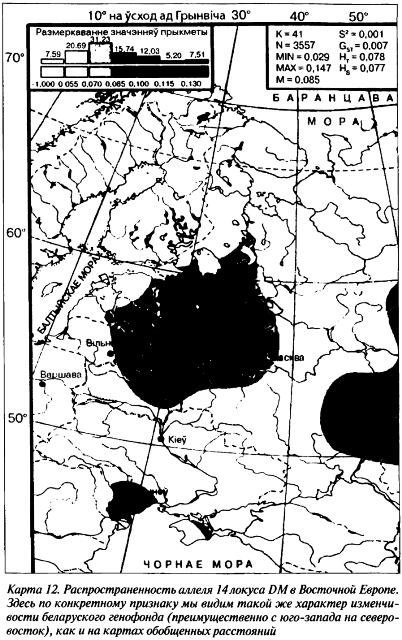
Распределение аллеля 14 и гетерозиготности по всему локусу DM на беларуской территории полностью соответствует ранее выделенному нами по антропогенетическим особенностям географическому направлению с юго-запада на северо-восток (карта 12). Наш суммарный анализ частот редких аллелей (от 20 и выше) показал, что их концентрация максимальна именно в беларуской популяции. Таким образом, этот пик, который до недавнего времени локализовали в Центральной Европе, переместился на территорию Беларуси. Этот факт еще раз демонстрирует необходимость и важность изучения антропологических и генетических признаков не только по их средним показателям, но и обязательно (как предлагает Ю. Г. Рычков) с учетом даже чрезвычайно редких признаков. То обстоятельство, что на частотный спектр аллелей локуса DM действительно влияют не только монголоидно-европеоидные соподчинения, но и естественный отбор (с последующим закреплением отдельных аллелей на конкретных территориях), достаточно аргументировано (Лимборская С. А. и др., 2002).
Наш предыдущий анализ полиморфизма триплетного повтора гена миотанинпротеинкиназы DM выявил 17 аллельных его вариантов с числом единиц от 5 до 29. Аллели с 6-м и 7-м повторами вообще не выявлены. Уровень гетерозиготности колеблется в разных популяциях от 71 до 81 процента, что статистически достоверно не отличает его от ожидаемого — 73–79 процентов (индекс фиксации — 0,002) и косвенно указывает на фактическую селекционную нейтральность нормального полиморфизма STG-повторов гена DM среди беларуских популяций.
Как видим, концентрации каждого из аллельных вариантов ДМ имеют свою особенность распространения среди коренных беларусов. Поэтому снова возникает потребность обращения к обобщенному массиву информации с помощью метода главных компонент частотных показателей посредством многомерного статистического анализа. Он помогает выявить общие закономерности изменчивости. Например, территория современной Беларуси находится в промежуточной зоне, где первая главная компонента определяет регион равновесия двух противоположных тенденций (Лимборская С. А. и др., 2002). В частности, северо-восточная и юго-западная провинции Беларуси имеют очевидную связь с соответствующими прилегающими территориями соседних стран.
Беларуские популяции изучены еще по одному микросателлитному локусу Cact685. Генетический анализ пока завершен в 4 выборках общей численностью 241 персона. Всего выявлены 12 аллельных вариантов СА-повтора. 16-й и 17-й повторы не выявлены вообще, кстати, как и у других представителей всей индоевропейской лингвистической семьи. По частоте заметно выделяются на гистограмме 19-й, 22-й и 27-й аллели — в пределах трети от всех наличных. Самые минорные аллельные повторы (20-й и 30-й) выявлены, соответственно, в гродненской сельской группе на северо-западе Беларуси и хойникской — на юго-востоке. Равновесие Харди-Вайнберга нарушено только в последней популяции. Этот факт косвенно может свидетельствовать о наличии здесь стрессовой экологической ситуации.
В качестве критерия внутрипопуляционного разнообразия избраны показатели ожидаемой (0,83–0,87) и наблюдаемой (0,82–0,88) гетерозиготности, что позволяет считать этот маркер высокоинформативным в популяционных и этногенетических исследованиях. Беларуский этнос соответствует своей европейской характеристике по мажорным концентрациям 22-го и 27-го аллелей в сравнении с народами уральской и алтайской лингвистических семей (Лимборская С. А. и др., 2002). На территории Беларуси их присутствие одинаково возрастает в направлении с юга на север (таблица 4).
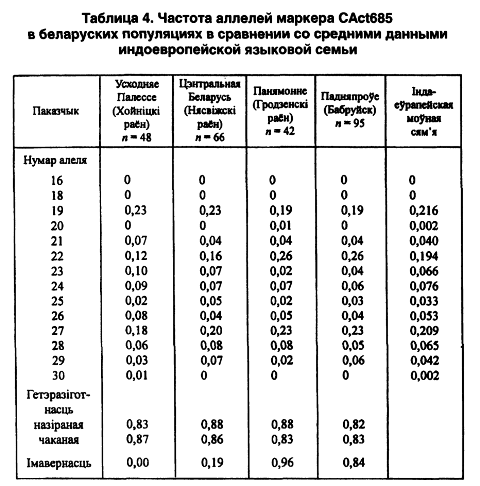
Однако 19-й аллель имеет противоположный вектор изменчивости. У минорных аллелей такая географическая зависимость не наблюдается (r = -0,59). По гетерозиготности локуса Cact685 регион ее минимальных показателей выявлен в Северной Беларуси.
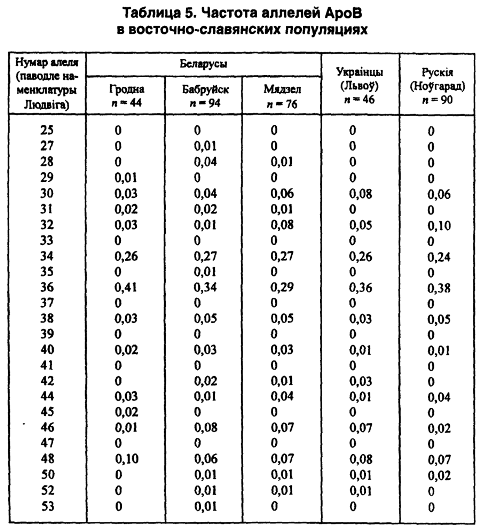
Кроме проанализированных выше микросателлитных маркеров, у части беларуских популяций изучались и отдельные мини-сателлиты, в частности, высокоизменчивый локус АроВ 2-й хромосомы. Последний характеризуется значительным полиморфизмом и обладает своими собственными популяционными отличиями. Всего среди этносов Европы выявлены 26 его аллельных вариантов, в том числе у беларусов 19 (в соответствии с номенклатурой Ludwig). Таким образом, здесь отсутствуют 7 аллелей — 25-й, 33-й, 37-й, 39-й, 41-й, 47-й, 55-й. Самые распространенные — аллели с повторами 34 и 36. Уровень гетерозиготности снова высокий, он составляет от 73 до 84 процентов. Распределение частот генотипов во всех популяциях отвечает равновесию Харди-Вайнберга.
У русских тоже нет 7 аллелей, но только три из них те же, что отсутствуют и у беларусов: 41-й, 47-й, 55-й. Эти редкие аллели отсутствуют у всех трех восточнославянских этносов. Среди украинцев нет почти половины евразийских аллельных вариантов — 12 (25,27, 28, 31,33, 37, 39,41,45,47, 53, 55). Таким образом, если у беларусов вместе с русскими одинаково отсутствуют только три аллеля, то у беларусов вместе с украинцами — уже семь, или в 2,3 раза больше. Этот факт свидетельствует о большей общности исторической судьбы нашего народа с южным соседом, чем с восточным.
Итак, в Беларуси явная геногеография конкретного признака обнаружена в самом мажорном 36-м повторе аллелей: от 0,29 в Нарочанском крае, через 0,34 в Бобруйском регионе до 0,41 — в Гродненском. Почти такая же геногеография сохраняется в случае частот аллелей 30 и 48 (таблица 5).
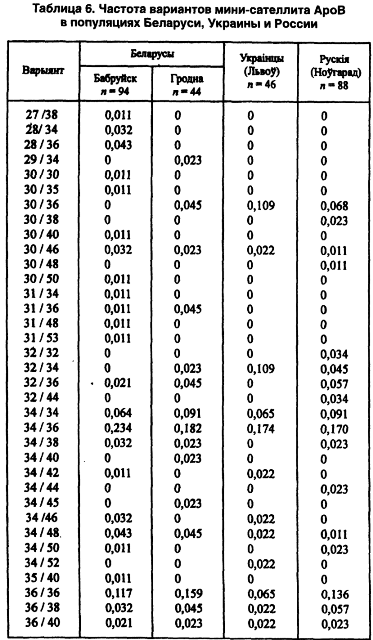
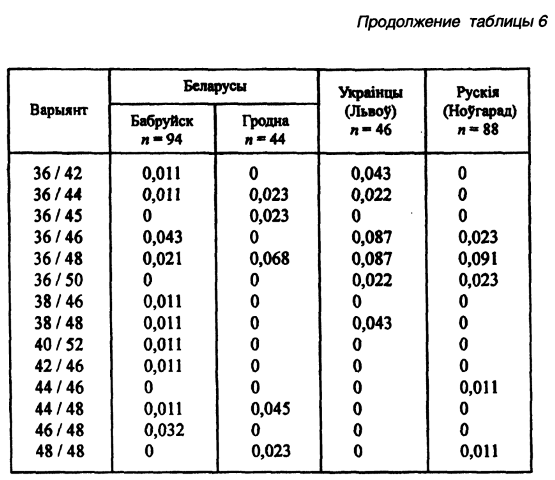
Другой модальный аллель (34-й) имеет одинаковую концентрацию во всех локальных популяциях — 0,26–0,27 (Лимборская С. А. и др., 2202). Мини-сателлит АроВ обладает выраженными расово-диагностическими свойствами. Согласно распределению его аллелей каждая из трех великих рас имеет свою особенность. Так, у европеоидов концентрация аллельных частот бимодальная, у монголоидов — унимодальная. На волжско-уральском пограничье встречаются популяции обеих этих рас. Гетерозиготность среди беларуских популяций не выходит за пределы 0,71–0,86, что полностью соответствует их европеоидности. Она не отличается от теоретически рассчитанной на статистически достоверном уровне. Все это, вместе взятое, способствовало широкому использованию данного признака в генетико-антропологических и этногенетических исследованиях (таблица 6).
Очередной маркер, полиморфизмом которого ныне широко пользуются геногеографы — мини- и микросателлитный мультилокусный ДНК-фингерпринтинг М13, называемый геномной дактилоскопией (по аналогии с классическим методом отпечатков пальцев). Большая часть вариантов минисателлита М13 отвечает критериям нормального полиморфизма. Поэтому данный метод востребован в современных популяционно-генетических исследованиях при анализе обобщенных биологических расстояний между популяциями разного иерархического уровня.
Нами по этому признаку пока изучены только популяции северо-запада и юго-востока Беларуси. С помощью компонентного и кластерного анализов в факторном пространстве действительно выявляется общее этническое «облако» (Шаброва Е. В., Микулич А. И., 2001). Анализ частотных характеристик в сравнении с финно-угорскими группами выявил значительную генетическую удаленность между ними. А по результатам многомерного шкалирования беларусы наиболее значительно отличаются от популяций коми.
Полиморфизм митохондриальной ДНК (мтДНК) в настоящее время тоже широко востребован антропогенетиками. В первую очередь потому, что она передается в наследство только по линии матери, а ее эволюция происходит за счет накопления мутаций в поколениях. Именно разнообразие митохондриального генофонда позволяет классифицировать молекулярные изменения в границах популяций и этносов. Так, если межпопуляционное различие по классическим маркерам составляет только 15 процентов от всей генетической изменчивости, то по мтДНК и генам Y-хромосомы — 30. Это весьма существенное различие, которое позволяет уточнить наши знания о геноме человека, приблизить открытие закономерностей в связях между популяциями и окружающим миром.
Наши совместные исследования с московскими и башкирскими коллегами показали, что индекс гаплотипового разнообразия мтДНК изменяется от 0,94 у беларусов до 0,96 у русских, 0,98 у башкиров и чувашей, 0,99 у татар и мордвы. Если учесть, что европейская раса характеризуется пониженной гетерогенностью, то наличие монгольского (или азиатского) компонента обозначается именно своей чрезвычайно высокой гетерогенностью по гаплотипам митохондриальной ДНК (таблица 7).
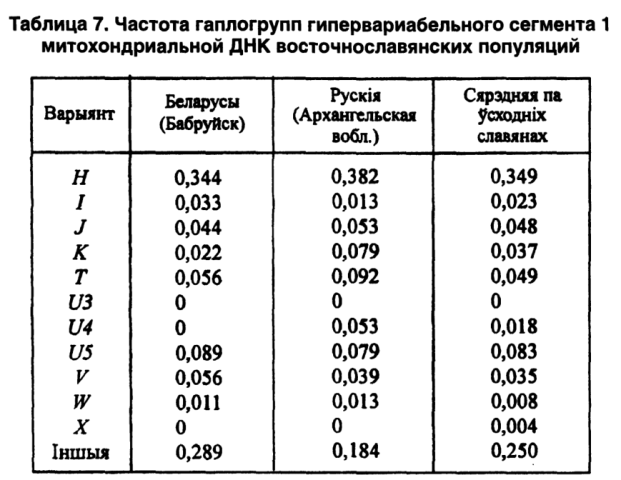
Среди исследованных восточнославянских популяций именно беларусы выделяются наличием гаплотипов 2, 12, 17, 22, 28. И только у нас отсутствуют 15 гаплотипов из 32 известных. По вариантам мтДНК (гаплогруппы Н, I, Т, U5, W, HV) беларусы приближаются к среднеевропейским популяциям (Лимборская С. А. и др., 2002).
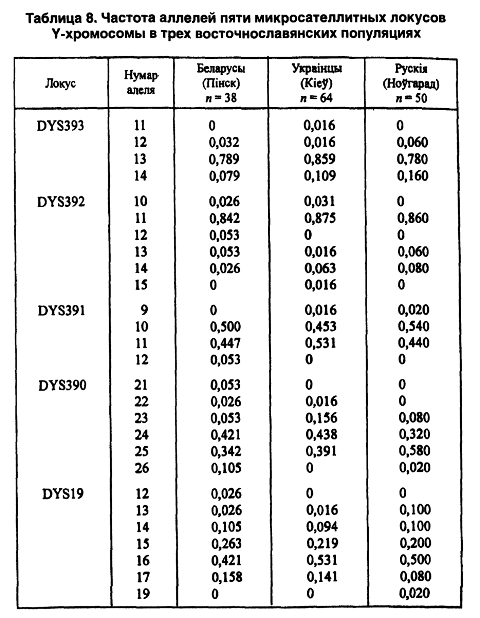
В настоящее время начато изучение полиморфных маркеров Y-хромосомы. Последняя находится в геноме мужчины и передается в наследство только по мужской линии. Y-хромосома обладает еще одной особенностью, значимой для антропологии, — чрезвычайно высокой географической и популяционной дифференциацией. Для анализа аллельного полиморфизма были выделены 5 микросателлитных локусов (DYS19, 390, 391, 392, 393) (таблица 8).
Индекс разнообразия подсчитали по формуле D = I - r2, где r — частота i-аллеля, или гаплотипа (Nei М., 1978). Достоверность различий в распределении частот аллельных вариантов и гаплотипов между обследованными популяциями определялась по Р. Фишеру (1958).
Среднее значение индекса разнообразия у беларусов Западного Полесья (0,52) статистически отличается от такового у русских и украинцев, где он составляет 0,47 и 0,45 соответственно. Наиболее значительная дифференциация восточнославянских этносов обусловлена локусом DYS390, но у беларусов еще в большей степени выделяется индекс аллельного разнообразия для локуса DYS19 (0,72).
В популяционных исследованиях локусы Y-хромосомы играют особенно важную роль. Анализ диаллельных маркеров показал, что все мужское население Европы сводится к 10 историческим родоводам (Semino О. Et al., 2000). Некоторые из них имеют достаточно древнее происхождение — от европейцев палеолита. Результаты нашего коллективного исследования восточных славян по распределению у них пяти аллельных вариантов показаны в таблице 8 (Кравченко С. А. и др., 2001). Индекс общего разнообразия среди трех этносов по всем изученным гаплотипам достаточно высок — 0,94. Только единичных гаплотипов зафиксировано 23 %, а гаплотипы 1-й и 2-й среди беларусов фактически доминируют — вместе около 24 %; 3-й, 5-й и 6-й образуют еще одну частотную группу — почти 19 %.
Несмотря на случайность популяционных выборок из восточнославянских этносов (Беларуское Полесье, Киев, Новгородчина), результаты генетического анализа позволили сравнить их между собой. Так, по двум позициям мажорных гаплотипов (1+2, 3+5+6) выделяются беларусы и украинцы. Однако среди последних значительно преобладает гаплотип 4 (12,5 %) — возможно, что он является для них этнически определяющим. У беларусов и украинцев одинаково отсутствуют 15-й и 16-й гаплотипы, а у русских и беларусов— 11-й, 12-й, 13-й.
По всем пяти локусам наибольшее число выявленных аллелей у беларусов — 23. Таким образом, из 27 аллелей среди беларусов не выявлены только 4 (по одному в каждом из пяти локусов, кроме DYS390); украинцы лишены 6 аллелей из 27, а русские — 7. Как видим, беларусы снова, аналогично случаю с системой резус (CDE), отличаются наличием редких аллелей. Это может свидетельствовать об их генетическом разнообразии и адаптационной пластичности к окружающей среде.
Существует масса доказательств наследственной обусловленности многих других особенностей биологии людских популяций. Например, наследственность иммунитета подтверждается передачей через поколения групп крови. Адаптационно значимым остается также биохимический полиморфизм, поскольку распределение частот генов и фенотипов зависит от комплекса экологических факторов (Спицин В. А., 1985). Проанализировав географию морфофизиологических комплексов в границах континентов, Т. И. Алексеева (1998) определяет эти комплексы как адаптивные типы и соглашается с вероятностью их наследственной природы.
Итак, научные данные свидетельствуют, что адаптивность формируется в результате длительной истории приспособления популяций человека — через множество поколений — к различным экологическим условиям существования. Использование экологического подхода к решению проблем медицинской генетики позволяет исследователям выделить новый научный раздел в антропологии — экогенетику.
В результате нашего наблюдения за генетическим статусом и состоянием здоровья городских и сельских популяций выявлен ряд кардио-респираторных патологий и значительное нейропсихогенное напряжение. Последнее истощает резервы функциональных возможностей человека, способствует их разбалансировке. При этом стрессовое состояние по своей биохимической основе не зависит от его первопричины — физиологической либо психологической (Новицкий А. А., Микулич А. И., 1994). Одно бесспорно, стрессовое воздействие комплекса субэкстремальных факторов ослабляет организм и формирует преморбидное[110] напряжение в ряде популяций, где процесс адаптации зависит от интенсивности и продолжительности отрицательного экологического воздействия.
Работа с новыми генокопиями предоставляет уникальную возможность использовать дополнительный антропогенетический полиморфизм в изучении адаптационной нормы современного населения применительно к экстремальным условиям существования. Феномен присутствия или отсутствия редко встречаемых аллелей почти всегда является внутренней характеристикой конкретной популяции. Отсюда возникает возможность отслеживания генетического гомеостаза, когда взаимоотношения между генами способствуют жизнеспособности организма в конкретных условиях внешней среды.
Из всего этого можно сделать следующие выводы.
1) Использование ДНК-маркеров в антропогенетических и геногеографических исследованиях значительно расширяет наши возможности в изучении этногенетических проблем происхождения конкретных популяций.
2) С увеличением количества этих маркеров, одинаково распределенных по геному человека, улучшается пренатальная диагностика наследственных патологий.
3) Медицинская генетика и антропогенетика приблизятся к выявлению причин злокачественных опухолей и их возможного предупреждения.
4) Генотипоскопия даже по 3–4 локусам, размещенным в разных хромосомах, позволит надежно идентифицировать геномы родителей и детей в случаях споров об отцовстве (материнстве).
5) Максимальное использование генома отдельного человека через призму популяционно-генетических характеристик позволит изучить влияние современной экосоциальной ситуации на возникновение новых патологий.
6) Высокая степень полиморфизма ДНК и обследование локальных популяций в географических пространствах радиусом 50–60 км, независимо от этнических и государственных границ, позволяют создавать географические карты антропогенетической и популяционно-демографической родственности.
Таким образом, мы еще раз убедились, что наряду с генетической причиной географической дифференциации популяций существует и экологическая.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК