Находки останков доисторических людей в Юго-Восточной Азии и Океании
Человек — единственный живой представитель семейства гоминид, так как человекообразные обезьяны относятся уже к другому семейству — понгид. Происхождение гоминид может быть прослежено от обезьяноподобного вида рамапитека (жившего 14 млн. лет назад), фрагменты челюстей которого найдены в Восточной Африке и на северо-западе Индии [1102; 129, с. 209]. Около 6 млн. лет назад в Южной и Восточной Африке появились представители рода австралопитековых. По-видимому, первые группы рода Homo, начавшие изготавливать орудия, происходили от одной из его ветвей. Точный ход эволюции человеческих предков в этот период еще неясен, но в Восточной Африке были найдены ранние останки гоминид, живших там около 2 млн. лет назад. А вместе с ними появились и первые грубые галечные орудия. Некоторые авторы называют этих самых ранних гоминид, изготовлявших орудия, Homo habilis. 1,5 млн. лет назад Homo habilis эволюционировали в Homo erectus, мозг которых был большего размера. К последним относятся самые ранние из ископаемых людей на Яве и в Китае, останки которых впервые были найдены в 1890 г.
В этом кратком обзоре важны два момента. Во-первых, поскольку древнейшие стадии человеческой эволюции зафиксированы лишь в Африке, она и представляется прародиной; откуда около 2 млн. лет назад человек начал расселяться в Азии. Разумеется, будущие открытия могут привести к пересмотру этой интерпретации, особенно если на Яве и в Китае будут встречены останки Homo habilis. Во-вторых, теперь уже общеизвестно, что обезьяны могут делать и в определенных условиях делают орудия, но человек на протяжении 2 млн. лет усложнял изготовление орудий. Ни одна обезьяна в диком состоянии не обработает даже галечное орудие (хотя в лабораторных условиях она способна на это), и африканский Homo habilis был, видимо, первым, кто перешагнул этот критический рубеж.
На Яве наиболее ранние гоминиды были найдены вместе с фауной джетис, относящейся к верхнему плиоцену или нижнему плейстоцену. Слой, в котором залегали фауна и останки гоминид, сформировался 3–1 млн. лет назад, но более точная их датировка неясна [819; 767]. Древнейшей находкой, возраст которой более 1,5 млн. лет, является череп ребенка из Моджокерто на Восточной Яве, отнесенный к виду Homo erectus. К несколько более позднему времени относятся три (а возможно, и четыре) обломка нижней челюсти особи с массивными челюстями, которую вначале назвали мегантроп яванский. Эти обломки были обнаружены на Центральной Яве, в Сангиране, причем по крайней мере один из них относится к верхней части слоя с фауной джетис Находка мегантропа усложняет проблему классификации.
Эволюция рода Homo происходила, видимо, без резких скачков. Это значит, что границы между Homo habilis и Homo erectus, а также между Homo erectus и Homo sapiens определяются больше договоренностью между специалистами, чем разрывами постепенности. Мегантропа, обладавшего массивными челюстями, сопоставляли с африканскими австралопитеками, а в 1964 г. П. Тобайяс и Г. фон Кенигсвальд сопоставили его с Homo habilis из Олдовайского ущелья в Танзании, возраст которого насчитывал 1,5–2 млн. лет [1376]. Но позже было высказано предположение, что эти челюсти, несмотря на массивность, укладываются в рамки вариативности яванского Homo erectus [906], чем дело и кончилось. Решить проблему можно будет только после новых открытий, однако следует отметить, что в Сангиране имелись и другие останки гоминид из верхней части слоя с фауной джетис, которые, несомненно, принадлежат к Homo erectus, так что к этой категории, возможно, относятся и все находки, связанные с фауной джетис.
Вместе с тем одно свидетельство позволяет предполагать появление гоминид на Яве еще до стадии Homo erectus. На верхней челюсти из Сангирана, относящейся ко времени джетис и, безусловно, к Homo erectus, сохранилась специфическая симиальная особенность в виде промежутка (диастемы) между каждым из верхних боковых резцов и клыками. У всех африканских австралопитеков диастема отсутствует, а поскольку она не является африканской чертой, можно предположить, что на Яве Homo erectus мог возникнуть независимо из австралопитековых Юго-Восточной Азии. Но пока что это лишь слабый намек на решение вопроса.
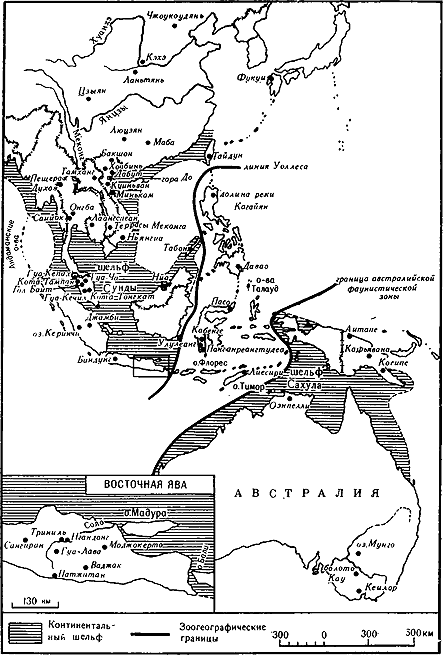
Юго-Восточная Азия с зоогеографическими границами и археологическими памятниками эпохи плейстоцена
На Яве большинство бесспорных находок Homo erectus происходят из отложений, содержащих фауну среднеплейстоценового комплекса триниль, имеющего возраст 1 млн. — 300 тыс. лет. К наиболее интересным останкам относятся почти целый череп из Сангирана (питекантроп 8), две черепные крышки, множество-зубов и других фрагментов [334; 149]. Для всей этой группы объем мозга варьирует в пределах от 750 до 1125 куб. см (включая находки зоны джетис), и, судя по реконструкции, ее признаками были вытянутый череп, наиболее широкий у основания, уплощение черепного свода, продольный гребень, массивные надглазничные валики, заглазничное сужение черепа, довольно прогнатное лицо с тяжелой нижней челюстью без подбородка. Зубной свод параболический, а зубы, особенно моляры, значительно крупнее, чем у современного человека, хотя морфологически с ними сходны.
В сравнении с современным человеком череп Homo erectus очень архаичен, но положение большого затылочного отверстия в основании черепа говорит о почти вертикальной осанке. В 1892 г. Е. Дюбуа обнаружил в Триниле бедренную кость современного облика с паталогическим разрастанием костного вещества (экзостоз). Многие годы считалось, что эта кость и несколько других фрагментов бедер принадлежали Homo erectus, и опубликованные в 1952 г. датировки по фтору как будто бы это подтверждали [105]. Однако последующие исследования заставили усомниться в происхождении этих костей [335]. Подозрителен хотя бы тот факт, что бедренные кости из Олдовайского ущелья в Танзании и из Чжоукоудяня под Пекином, принадлежащие, безусловно, Homo erectus, отличаются гораздо большей архаичностью. Но если даже проигнорировать яванское бедро, все равно останется несомненным, что и австралопитеки, и Homo erectus были почти прямоходящими существами. Безусловно, эволюция черепа до этого времени шла в-ином, замедленном темпе в отличие от эволюции осанки и зубной системы.
Другим интересным фактом, связанным с яванским человеком, является то, что он или его материковые родственники могли жить одновременно с гигантскими (трехметрового роста) понгидами, названными Gigantopithecus blacki [1260]. Первые останки гигантопитека были обнаружены в Гонконге, а в 1956–1958 гг. на юге Гуанси-Чжуанского автономного района в Китае было найдено более 1 тыс. зубов и три нижние челюсти. Останков пост-краниального скелета пока не обнаружено, однако огромные зубы указывают на человекообразную обезьяну, питавшуюся семенами растений и обитавшую в открытой местности в Юго-Восточной Азии в период между плиоценом и средним плейстоценом. В 1946 г. Ф. Вайденрайх опубликовал интересную книгу, в которой высказал предположение, что эти гиганты являлись предками человека и он развился из них через ряд промежуточных — форм уменьшающихся размеров, представленных мегантропами и питекантропами (Homo erectus) [1437]. Некоторые гоминидные черты в зубной системе гигантопитека говорят о том, что в глубочайшей древности линии эволюции его предков и предков человека могли быть связаны, но сам он представлял, как теперь видно, высокоспециализированный вид, полностью вымерший.
Ископаемые останки доисторических людей в Китае в целом аналогичны яванским. В Ланьтяне, в провинции Шэньси, в 1963 г. обнаружена нижняя челюсть, а в 1964 г. — черепная коробка [1472; 4]. Черепная коробка столь же архаична, как находки периода джетис с Явы, и ее объем равен 780 куб. см. Вместе с ней были найдены орудия из отщепов и останки фауны нижнего или начала среднего плейстоцена. Поэтому возраст черепной коробки может достигать 700 тыс. лет и более. Нижняя челюсть, видимо, моложе, ее возраст — около 300 тыс. лет[3]. На ней отсутствует третий моляр, что сейчас характерно для 30 % монголоидов [757].
Наиболее известные находки Homo erectus (или синантропа) были сделаны начиная с 1921 г. в нижней пещере (местонахождение 1) Чжоукоудянь, в 42 км юго-западнее Пекина. Большинство этих останков во время второй мировой войны было утрачено, но, к счастью, с них успели сделать слепки. Основные находки — это 14 черепов различной степени сохранности, 12 нижних челюстей со множеством утерянных зубов и несколько костей посткраниального скелета. Эти формы прогрессивнее яванских. Объем их черепа варьирует от 850 до 1300 куб. см, составляя в среднем 1075 куб. см [871, с. 105]. Черепа еще имеют большую ширину основания, но свод черепов выше и более округлый, чем у яванских людей, а надглазничные валики и зубы меньших размеров. В Чжоукоудяне были найдены семь бедренных костей, две плечевые и одна ключица. Их сходство с современными формами говорит о почти вертикальном положении тела при хождении. Таким образом, по физическим особенностям местное население было более развитым, чем яванское, и, судя по фаунистическим и пыльцевым данным, обитало в лесостепи с умеренным климатом, что, видимо, коррелирует со вторым интерстадиалом в Гималаях [756; 830]. Последний не имеет четких дат, но, по оценке, возраст пекинского человека составляет 300–200 тыс. лет [1030; 222, с. 50][4].
На Яве имеется лишь одна группа останков, заполняющая лакуну между Homo erectus и современным человеком. Это 11 черепных крышек и две большие берцовые кости, найденные в 1931–1933 гг. на предположительно верхнеплейстоценовой террасе на р. Соло в Нгандонге в центральной части Явы. Найденная с этими останками фауна состоит из 25 тыс. костей млекопитающих, в том числе пантеры, носорога, гиппопотама, кабана, оленя, буйвола, а также современного и ископаемого слонов. Все это предполагает степную среду. Четкой датировки местонахождения пет; предположение о верхнеплейстоценовом облике фауны не меняет дела. Т. Джекоб определяет возраст находки в 100 и 60 тыс. лет [766, с. 39]. Лицевой скелет и основания черепов отсутствуют, поэтому некоторые авторы видят в солоском и пекинском людях каннибалов. Изучив черепа людей, подвергшихся каннибализму, из Новой Гвинеи, Джекоб довольно убедительно оспаривает положение о том, что солоский человек был каннибалом; Г. фон Кенигсвальд первоначально предполагал, что солоские черепа могли служить чашами [1438; 768].
Многие современные авторы рассматривают солоского человека как развитую форму Homo erectus, лишь немногим более архаичную, чем европейские неандертальцы. Промежуточное положение солоского человека привело к терминологическим спорам, его называют то Homo erectus soloensis, то Homo sapiens soloensis, то Pithecanthropus soloensis. К. Кун считает, что средний объем мужских черепов солоского человека составлял 1150 куб. см. Кун относит солоского человека к Homo erectus, т. е. к тому же этапу эволюции, что и пекинского человека, хотя первый датируется более поздним временем. Я, основываясь на интуиции, склонен считать, что солоский человек жил, как минимум, 100 тыс. лет назад, но, возможно, и гораздо раньше. Так как в настоящий момент эта проблема неразрешима, можно лишь прибегнуть к гипотезам. Две из них заслуживают особого внимания. В соответствии с первой, древность солоского человека — не более 60 тыс. лет. В этом случае он скорее всего представлял вымершую ветвь [129, с. 319], так как люди современного вида жили в Индонезии уже 40 тыс. лет назад, а 20 тыс. лет слишком короткий период для эволюции на месте. Вымереть могли и более ранние представители рода Homo erectus на Яве, но тогда неясно, как эта линия соотносилась с другими линиями развития в других местах. Согласно второй гипотезе, которая кажется довольно правдоподобной, солоский человек, будучи гораздо более древним, мог являться одним из прямых предков современного человека, в особенности австралоидов в Юго-Восточной Азии. Так как в плейстоцене Ява была островом, местное население, находившееся в изоляции, конечно, могло иметь много архаических черт. Но маловероятно, что оно вымерло, не оставив потомков. Видимо, современные австралоиды Юго-Восточной Азии до некоторой степени — потомки солоской линии, в которую вливалось достаточно материковых генов, чтобы она избежала превращения в особый вид.
Homo erectus и человек современного вида в Восточной Азии
Чтобы отличить современного человека от более архаичных представителей того же вида, таких, как Homo sapiens neanderthalensis и Homo sapiens soloensis, его классифицируют как подвид Homo sapiens sapiens. Древнейшие ископаемые останки человека современного вида имеют возраст 60 тыс. лет, а 40 тыс. лет назад он уже широко распространен в Старом Свете. Происхождение человека современного вида неясно; неандертальские и солоские формы обычно не считаются его основными предками, хотя некоторыми из своих генов он, несомненно, обязан им. Развитие Homo sapiens от Homo erectus началось, видимо, 200–300 тыс. лет назад, когда объем мозга превысил 1300 куб. см, но исследователям об этом ничего не известно[5].
Попробуем проанализировать, как Homo erectus соотносится с современными австралоидами и монголоидами. Гипотеза о частичной преемственности кажется более приемлемой, чем гипотеза о полной смене, потому что частичная преемственность реальнее полного вымирания более ранних форм. В пользу первой гипотезы говорят некоторые палеоантропологические данные, о чем речь пойдет ниже.
Монголоиды Восточной Азии и Америки — широко распространенная и очень вариативная группа. Антропологические признаки, объединяющие большинство монголоидных народов, — это темные глаза, прямые волосы и широкое, плоское лицо [757, с. 2]. Эти признаки выработались, видимо, на протяжении длительного времени в результате естественного отбора среди довольно вариативного населения, обитавшего где-то в Восточной Азии. Уже говорилось о том, что в желтоватой коже монголоидов содержится мало меланина, но зато эпидермис насыщен кератином. Такая кожа хорошо отражает прямой солнечный свет и препятствует проникновению ультрафиолетовой радиации в большей мере, чем кожа европейцев [280, с. 234]. Однако Восточная Азия, за исключением территории, примыкающей к Тонкинскому заливу, не является районом высокой солнечной радиации. Поэтому сопротивляемость ультрафиолетовому излучению не могла быть единственным действующим фактором отбора. К. Брейс предложил другую причину относительной светлокожести монголоидов. По его мнению, она была вызвана употреблением одежды в северных широтах во время последнего оледенения, что снизило адаптивное значение темной кожи [280, с. 219, 148]. По современным палеоантропологическим данным, вероятный район происхождения монголоидов — Северный и Центральный Китай, где употребление одежды могло возникнуть очень давно.
Конечно, очень трудно проследить происхождение монголоидов по антропологическим останкам. Однако в ископаемых материалах можно выявить несколько характерных особенностей. Это лопатообразность верхних резцов, врожденное отсутствие третьих моляров, подбородочный бугор, сагиттальный валик, кость инков, широкие носовые кости и платимерия (относительная уплощенность диафиза бедра) [757, с. 3]. Далеко не у всех монголоидов есть эти особенности, к тому же и они свойственны не только монголоидам. Таким образом, можно говорить только о тенденции: у монголоидов эти черты встречаются чаще, чем у представителей других рас.
Ф. Вайденрайх, хорошо знакомый с палеоантропологическими материалами, вначале предполагал, что по крайней мере некоторые группы монголоидов были прямыми потомками пекинского человека, о чем, в частности, свидетельствовали лопатообразные резцы и подбородочный бугор [1437, с. 84]. К. Кун решил, что нашел подтверждение этому предположению, когда в 1962 г. насчитал 17 общих для этих групп антропологических особенностей [279]. Он заявил также, что смог проследить эволюцию монголоидов от пекинского человека через серию средне- и верхнеплейстоценовых находок в Китае. Ранние формы человека современного монголоидного вида представлены верхнеплейстоценовыми черепами из Цзыяна в провинции Сычуань и Люцзяна в Гуанси-Чжуанском автономном районе [1471; 216, с. 752–754]. Обе эти находки плохо датированы, но Д. Хьюз согласился с предположением К. Куна о том, что они представляют начало выделения монголоидной расы [757, с. 7]. Более поздние останки человека в Китае, например из верхней пещеры Чжоукоудянь, все нынешние авторы связывают с монголоидами[6].
Эти данные, конечно, довольно уязвимы, особенно когда речь идет о датировках, но все же сейчас можно, видимо, говорить, что современные монголоиды по крайней мере в определенной степени происходят от пекинского человека. Нет никаких данных в пользу предположения о его истреблении и смене какими-либо мигрантами современного вида. Однако нельзя утверждать и того, что пекинский человек был главным источником эволюции монголоидов. Видимо, антропологическая ситуация в Восточной Азии в среднем или позднем плейстоцене была столь же разнообразна, как и сейчас, и многие гетерогенные популяции передали свои гены нынешней гетерогенной группе монголоидов. «Если бы даже мы могли реконструировать запутанную картину сменяемости смешанных групп, участвовавших в формировании современных популяций, в окончательном виде она скорее напоминала бы сетку, чем дерево», — пишет Ж. Йерно [989, с. 42].
Если обратиться к Юго-Восточной Азии, то картина будет еще более сложной, чем в Китае, так как здесь монголоиды и австралоиды долгое время развивались бок о бок и гибридизировались. В районах от Центрального Китая до Юго-Восточной Азии не было существенных барьеров для локальных передвижений в меридиональном направлении, и, видимо, человек современного вида формировался здесь в обстановке клинальной изменчивости. Возможно, на севере шла аккумуляция монголоидных черт, а на юге — австралоидных, но вряд ли между этими Двумя регионами пролегала резкая граница.
Зная современное распространение фенотипов в Юго-Восточной Азии, Западной Меланезии и Австралии, проще всего предположить, что монголоиды двинулись на юг и сменили там австралоидов, которые первоначально занимали территорию от Юго-Восточной Азии до Новой Гвинеи и Австралии, а теперь сохранились в Юго-Восточной Азии лишь в виде небольших изолированных групп негритосов. Но к подобным идеям следует относиться с осторожностью, учитывая сложную ситуацию обмена генами между двумя весьма вариативными расами, которые могли входить в единый ареал клинальной изменчивости. Было бы наивно предполагать массовую миграцию однородного монголоидного населения, истребившего своих австралоидных предшественников.
Гипотеза Ф. Вайденрайха, поддержанная К. Куном, сводится к следующему: австралоиды формировались параллельно монголоидам и прямо происходили от юго-восточноазиатской популяции Homo erectus, представленной яванскими находками. Эта линия эволюции включала солоского человека и две плохо документированные находки останков черепов, сделанные Е. Дюбуа в 1890 г. в Ваджаке на Центральной Яве. Эти черепа, относящиеся к верхнему плейстоцену или голоцену [766, с. 51], принадлежали Homo sapiens sapiens, хотя один череп сохранил довольно выраженные надглазничные валики и до некоторой степени развитой сагиттальный валик. Кроме того, имеется известный череп Homo sapiens sapiens из пещеры Ниа в Сараваке, который Д. Бросуелл сопоставлял с тасманийскими черепами [157]. Полагают, что древность этого черепа — 40 тыс. лет, но есть основания сомневаться в правильности такой датировки. Если же она верна, то череп из Ниа принадлежал древнейшему из известных сейчас австралоидов.
В настоящий момент гипотеза Вайденрайха — Куна о непрерывном развитии в Юго-Восточной Азии кажется наиболее приемлемой для объяснения эволюции австралоидов. Но недавние открытия в Австралии снова подняли вопрос о смене населения, пусть только на локальном уровне. Древнейшие останки людей в Австралии представлены кремацией женщины на стоянке у оз. Мунго на западе Нового Южного Уэльса [146; 147][7]. Нет оснований сомневаться, что у Мунго найдены останки прямого предка современных аборигенов. Однако в связи с открытием нескольких погребений людей очень грубого архаичного типа в болоте Кау в Северной Виктории [1370; 1371] возникли новые вопросы. Эти люди жили 10 тыс. лет назад и, таким образом, синхронны австралоидам типа Мунго, но они сохраняют так много особенностей Homo erectus, что не могут относиться к населению тога же физического облика. Нижние челюсти у некоторых из них еще массивнее, чем у Homo erectus с Явы, а толщина черепного свода достигает 2 см. По общему облику они не выходят за рамки вариативности Homo sapiens, но их, безусловно, нельзя объединять с современными австралоидами.
Хотя находки в болоте Кау относятся к довольно позднему времени, они свидетельствуют о возможности очень раннего заселения Австралии солоскими людьми, которые позже на протяжении длительного периода были сменены и частично поглощены пришелицами-австралоидами[8]. Но и в этом случае трудна объяснить, почему первые сохранились в течение 15 тыс. лет — ведь женщина из Мунго жила 25 тыс. лет назад, а самое позднее из погребений в болоте Кау имеет древность всего в 9 тыс. лет. Австралия остается за пределами этой книги, поэтому здесь достаточно отметить, что она была заселена австралоидами по крайней мере 25 тыс. лет назад[9], позже здесь могла произойти смена населения. Теории заселения Австралии актуальны вообще, а в последнее время в особенности. Находки в болоте Кау, безусловно, доказывают, что заселение Австралии происходило неоднократно, однако скольким именно миграциям аборигены обязаны вариативностью своего современного антропологического типа, остается неясным [128; 1369; 1754].
Однако вернемся к австралоидам Юго-Восточной Азии конца плейстоцена и голоцена. Картина очень осложнена из-за малочисленности раскопок и отсутствия датировок. Преобладает мнение, что одна из главных волн распространения монголоидного фенотипа, в Индонезии и на Филиппинах в особенности, восходит к неолитическому времени [1171; 815]. По современным данным, это началось до 3000 г. до н. э. Донеолитическое население Индонезии и Филиппин было, видимо, в основном австралоидным, а создатели донеолитического хоабиньского технокомплекса на материковой части Юго-Восточной Азии обладали, очевидно, смешанными австралоидно-монголоидными чертами. Эта упрощенная схема, возможно, сильно отличается от реальной, но пока лучше ничего нельзя предложить. Во всяком случае, ясно, что речь не может идти о быстром «поглощении» австралоидов монголоидами.
По мнению К. Куна, уже к началу неолита в Юго-Восточной Азии распространилось значительное число монголоидных генов. Но он все же считал, что основная волна монголоидов распространилась в результате ханьской экспансии на рубеже новой эры [279, с. 416]. В таком случае еще 2000 лет назад в материковой части Юго-Восточной Азии могла сохраниться довольно крупная австралоидная популяция. По-видимому, так оно и было. Даже в китайском описании населения индианизированного царства Фунань в Камбодже (250 г. н. э.) содержится много упоминаний о темной коже и вьющихся волосах [1043, с. 254], хотя современное население Кампучии почти целиком относится к монголоидному фенотипу. Приведенная реконструкция опирается на малочисленные палеоантропологические данные, но в целом вырисовывается довольно связная картина. Многие скелеты хоабиньского времени из Северного Вьетнама, Таиланда и с Малаккского полуострова имеют смешанный монголоидно-австралоидный облик [947; 946; 378; 1181, с. 31; 1070; 1166; 1395, с. 16–18; 1383; 766]. По-видимому, речь может идти о клинальной изменчивости с вероятным преобладанием австралоидных черт на полуострове. Однако, за исключением факта появления монголоидного населения в Западном Таиланде к 2000 г. до н. э. [1166], у нас почти нет палеоантропологических данных о монголоидной экспансии на материке в неолите. Таким образом, остается вернуться к предположениям, высказанным в предыдущем абзаце.
В Индонезии и на Филиппинах монголоиды распространились в ареале, который прежде был австралоидным. Выше уже упоминался австралоидный череп из пещеры Ниа в Сараваке. Помимо него в той же пещере было найдено несколько донеолитических погребений, возраст которых — 17—6 тыс. лет; зубная система останков отличалась «меланезоидными» чертами [817; 666; 648]. Погребенные из вышележащих неолитических слоев имели уже монголоидную зубную систему. Они датируются концом II–I тысячелетием до н. э.
Т. Джекоб детально проанализировал много палеоантропологических останков с о-вов Ява, Сулавеси и Флорес [766], хотя только некоторые из них были увязаны с надежным археологическим контекстом. К меланезийскому и австромеланезийскому типу Джекоб отнес скелеты и другие останки со стоянок Восточной Явы и Флореса. Стоянки относятся, видимо, к раннему неолиту, а палеоантропологические останки в целом — к австралоидной группе. Методом радиоуглеродного анализа было установлено, что возраст одной из стоянок Флореса — 1600 лет до н. э., но это не решает вопроса о монголоидной волне, ибо на Флоресе до сих пор обитает население с австралоидными чертами. Большее значение имеют находки в пещере Гуа-Лава около Сампунга на Яве, так как в этом районе сейчас преобладают монголоиды, однако отсутствие датировки не позволяет делать какие-либо выводы. Зато мы можем дополнить наши данные серией из 2682 зубов, найденных в пещере Леангкаданг на юге Сулавеси. Джекоб назвал их безусловно монголоидными. В Леангкаданге был найден комплекс тоалских отщепов и пластин, которые датируются примерно 5000 г. до н. э., однако, учитывая аналогичные находки с других местных стоянок, можно предположить, что зубы относятся к периоду не ранее I тысячелетия до н. э. Данные Т. Джекоба позволяют считать, что в донеолитическую эпоху индонезийское население в своей основе было австралоидным, но уже в ранние периоды голоцена, вероятно, через Филиппины сюда начали проникать монголоиды.
На Филиппинах древнейшие человеческие останки представлены передней частью черепа, найденной в одной из пещер Та-бон на Палаване, заселенной 22–24 тыс. лет назад [475, с. 40–44]. Данные об этой находке полностью еще не опубликованы. В том же слое позже была найдена нижняя челюсть, которую Н. Макинтош назвал австралоидной [925, L]. На Палаване в пещере Дуйонг обнаружено неолитическое погребение, относящееся к началу III тысячелетия до н. э. [475, с. 60], но и эти палеоантропологические данные еще не опубликованы. В пещерах Табон удалось вскрыть серию погребений в сосудах, относящихся ко второй половине II тысячелетия и I тысячелетию до н. э. По ярко выраженной скуластости и лопатообразным резцам Н. Уинтерс определил их как монголоидные [1486][10].
Таким образом, в островной части Юго-Восточной Азии останки монголоидов доисторической эпохи пока найдены только на Филиппинах, Сулавеси и в Сараваке. Конечно, по этим случайным находкам трудно судить о реальной картине. Однако описанные материалы хорошо увязываются с лингвистическими и археологическими данными, свидетельствующими о том, что предки полинезийцев и микронезийцев пришли с Филиппин или из северо-восточной части Индонезии примерно 4 тыс. лет назад. В их жилах текла преимущественно монголоидная кровь.
Итак, данные, полученные на островной части Юго-Восточной Азии, могут свидетельствовать о длительном проживании австралоидов на о-вах Сунды по крайней мере до 1000 г. до н. э. или еще позже. Однако к 3000 г. до н. э. или еще раньше сюда начали проникать монголоиды, пришедшие через Филиппины на север Калимантана и на Сулавеси. Последняя значительная волна монголоидов, вполне вероятно, могла распространиться на западе Индонезии за последние 2500 лет. Что касается причины распространения монголоидов в островной части Юго-Восточной Азии, то предположительно его можно связывать с общим ростом народонаселения в неолите вследствие развития земледелия и оседлости. Более поздние хозяйственные успехи индианизированных царств, несомненно, ускорили этот Процесс.
Антропологическая история Океании
Новая Гвинея была заселена более 30 тыс. лет назад, но до сих пор там не обнаружено человеческих останков эпохи плейстоцена. Генетические и фенотипические данные позволяют предполагать, что население Новой Гвинеи находится в тесном родстве с австралийскими аборигенами. Но если австралийские аборигены развивались в длительной изоляции, то папуасы смешивались с людьми, имевшими монголоидные черты и заселившими Меланезию, Микронезию и Полинезию в последние 5 тыс. лет.
Таким образом, в истории формирования антропологического типа в Меланезии можно выделить два основных этапа. Более длительный первый этап, начавшийся 30 тыс. лет назад, — это заселение Новой Гвинеи и соседних островов и первоначальное развитие здесь австралоидов. Второй этап, который охватывает последние 5 тыс. лет, совпал с распространением австронезийских языков и расселением клинальных групп с более выраженным монголоидным фенотипом. Эти группы отличались от классических материковых монголоидов и скорее были переходным типом (встречается сейчас на востоке Индонезии).
В целом эта эволюционная модель основывается на идеях К. Куна. Но в прошлом высказывались иные точки зрения. В 1937 г. У. Хауэлле попытался объяснить разнообразие антропологических типов в Меланезии четырьмя миграциями. Первыми были австралийские аборигены, заселившие Новую Гвинею, о-ва Бисмарка и Новую Каледонию. За ними пришли негритосы, затем негры, последними были небольшие миграции из Полинезии и Микронезии. В 1949 г. Дж. Бердселл выдвинул свою теорию, сходную с гипотезой Хауэллса. По мнению Бердселла, первыми пришли негритосы, с которыми связаны многие черты папуасов, обитающих в Нагорьях Новой Гвинеи. За ними последовали две другие группы, названные исследователем муррайями (архаические кавказоиды) и Карпентариями (австралоиды). Большого влияния на Новую Гвинею они не оказали, но муррайи оставили глубокий след в истории севера Новой Каледонии. Последняя, поздняя группа состояла из монголоидов. И Хауэлле, и Бердселл считали, что антропологическая картина в Меланезии — результат гибридизации этих четырех групп.
Хауэлле и Бердселл были единодушны в том, что монголоиды пришли последними, но в вопросе о первых трех миграциях их мнения разошлись. Однако ни то, ни другое не подтверждается современными данными. Если не пытаться отделить негритос-скую миграцию от австралоидной, то почти нет оснований говорить о более чем двух миграциях. Некоторые антропологи, негативно относящиеся к теориям миграционизма, впали в другую крайность. «Современное меланезийское население является продуктом длительных сложных изменений, — пишет Д. Суиндлер, — причем его крайнее разнообразие, так бросающееся в глаза, было вызвано в первую очередь такими процессами, как мутация, миграция, естественный отбор, генетический дрейф и предпочтительные браки… Первичные компоненты, позже смешавшиеся, могли быть представлены, многочисленными разнотипными группами, медленно проникавшими в Меланезию из Азии и заселявшими местные острова. Они смешивались с некоторыми соседними группами, но по отношению к другим сохраняли эндогамию. Что представлял собой этот процесс: три-четыре отдельные миграции или медленную инфильтрацию различных мелких групп? Последнее кажется более правдоподобным» [1352, с. 48–49].
Суиндлер не прав, отказываясь от идеи нескольких миграций. Археологические и лингвистические данные позволяют предполагать, что в Меланезии наблюдалось два основных периода миграций, а в промежутке между ними могла происходить инфильтрация мелких групп. Этими двумя крупными группами были австралоиды и более монголоидное население. В Меланезии это население широко смешивалось с ранее пришедшими австралоидами, но в Полинезии и Микронезии оно сохранило свой облик. Две последние области были заселены исключительно австронезийцами, носителями значительного монголоидного наследия.
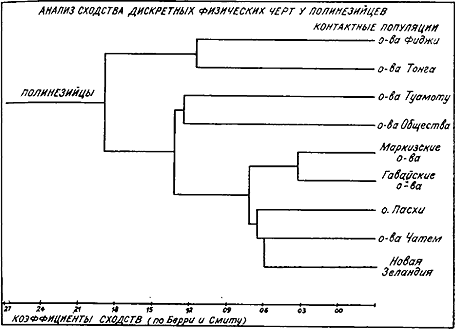
Сходство полинезийцев по антропологическим признакам (по М. Петрусевски)
Сколько миграций было необходимо для возникновения современной картины в Полинезии и Микронезии? В Полинезии, как показывают археологические и лингвистические исследования, была лишь одна крупная миграция, в Микронезии, видимо, две: одна с запада, другая с юга, но их нельзя расчленить исходя из современного антропологического типа, распространенного здесь. Вот почти все, что можно пока сказать о Микронезии. Что касается Полинезии, то ранние авторы писали о миграциях двух и более отдельных групп. Так, в 1924 г. Л. Салливен высказал мысль о четырех компонентах формирования полинезийцев: двух кавказоидных, одном негромеланезоидном и одном негромонголоидном. В прошлом представители этих антропологических типов в разной пропорции комбинировались на разных островах. В 1943 г. Г. Шапиро высказал предположение, что вначале мигрировало долихокефальное население, которое в окраинных районах Полинезии сохранялось вплоть до появления европейцев, а затем происходила миграция брахикефального населения, ставшего основным в Центральной Полинезии и на Гавайях. Выводы Г. Шапиро хорошо аргументированы и до сих пор не потеряли своей значимости. «В основе полинезийского населения, — писал он, — лежит единый антропологический тип, что свидетельствует о происхождении последовательных волн пришельцев из одного источника. Принадлежность разных волн мигрантов к различным антропологическим типам представляется весьма сомнительной» [1208]. Этот вывод приемлем и сейчас, однако мнение о том, что в Полинезии наблюдалось более одной миграции, теперь можно поставить под сомнение, хотя на основе одних только антропологических данных опровергнуть его невозможно.
Недавно М. Петрусевски, используя многомерные статистические методы, провел исследование множества фиджийских и полинезийских черепов и выделил три основные группы. В первую вошли черепа с Фиджи, Тонга и Самоа, для которых характерны промежуточные (меланезийско-полинезийские) черты, во
вторую — черепа с о-вов Общества и Туамоту (Центральная Полинезия), а также, видимо, с Маркизских островов, в третью — черепа с Гавайских островов, Новой Зеландии, о-вов Чатем, Пасхи (Окраинная Полинезия) и, возможно, с Маркизских островов. Место последних в этой классификации определяется применяемой статистической методикой, но в целом группы различаются довольно четко, однако эти различия вовсе не обязательно связаны с отдельными миграциями извне. Более правдоподобно предположение о локальной дифференциации, сочетавшейся с небольшими передвижениями населения.
Выводы Петрусевского не неожиданны, так как в течение длительного периода между фиджийцами, тонганцами и самоанцами иногда заключались браки, как и между островитянами о-вов Общества и Туамоту. Острова Окраинной Полинезии были более изолированы, и, возможно, население сохранило черты общего древнего полинезийского фенотипа. Что касается происхождения полинезийцев в целом, то лингвистами и археологами сейчас доказано, что полинезийский треугольник был заселен выходцами из одной или нескольких изолированных общин, обитавших скорее всего где-то в районе Тонга или Футуны. Это произошло на рубеже II–I тысячелетий до н. э. или чуть ранее. Ниже мы остановимся на этом подробнее, однако следует подчеркнуть, что данные антропологии и генетики не противоречат гипотезе происхождения полинезийцев от единого, монголоидного в своей основе населения, которое в предшествующий период имело в Меланезии контакты с австралоидами. Как отмечал К. Кун, «полинезийцы являются такой же частью монголоидно-австралоидной клинальной цепи, как формозцы, филиппинцы и балийцы, но у них отмечается больше австралоидности, чем у некоторых из последних» [280, с. 184]. То же относится и к микронезийцам. Теория об одной основной миграции в Полинезию не исключает вероятности нерегулярного прилива генов из Америки. Однако трудно избавиться от впечатления, что Полинезия заселялась только единожды и заселение шло с запада.
Как случилось, что полинезийцы и восточные микронезийцы достигли своих островов, не оказав более существенного влияния на Западную и Центральную Меланезию и, в свою очередь, избежав более сильного воздействия оттуда? Для ответа на этот вопрос следует привлекать данные не только антропологии, но и других наук, о чем речь пойдет ниже. Здесь же отметим, что полинезийцы и восточные микронезийцы распространялись через Меланезию и не входили в очень тесные генетические контакты с меланезийцами, которые ко II тысячелетию до н. э. уже расселились на восток вплоть до Новых Гебрид и Новой Каледонии, Переселившиеся в Полинезию и Микронезию австронезийские по языку группы в основном сохранили свою монголоидность, а те, которые остались в Меланезии, частично, но не полностью смешались с окружающими меланезийцами.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК