Глава 10 ДНК-генеалогия Белоруссии и ее сходство с ДНК-генеалогией России
Изучение истории славянских популяций интересно и с научной, и с общепознавательной точки зрения. Оно активно продолжается последние 200 лет, а то и дольше, принимая во внимание труды и полемику М.В. Ломоносова с его немецкими (в основном) оппонентами. Несмотря на убеждение М.В. Ломоносова: «Что славенский народ был в нынешних российских пределах еще прежде Рождества Христова, то неоспоримо доказать можно» (Ломоносов М. В., Пол. собр. соч. Т. 6. Труды по русской истории, общественно-экономическим вопросам и географии. 1747–1765. – М.-Л.: АН СССР, 1952. С. 22), высказанное им в известной дискуссии с профессором Г.Ф. Миллером, «немецкая школа» в академических кругах фактически (пока) одержала верх, и в академической науке продолжают считать, что славяне пришли на занимаемые ими сейчас земли в середине 1 тыс. нашей эры. Откуда пришли, где жили их предки до середины 1 тыс., и «прежде Рождества Христова», на каких языках разговаривали – академическая наука ответы пока не сформулировала.
Откуда вообще могут появиться новые убедительные, объективные данные? В этом отношении имелись всего три принципиальных источника информации – археология, письменные материалы (изучение старинных фолиантов, включая легенды-сказания-мифы и др.), лингвистические сведения (включая топонимику, ономастику и т. п.). Принципиально важные сведения из перечисленных источников уже получены предыдущим развитием наук, однако их интерпретация в наше время также приобрела дискуссионный характер.
Научный прогресс предложил новый путь – ДНК-генеалогию, «молекулярную историю». Получаемые данные являются объективными. Они формируются в виде мутаций в ДНК, что дает возможность оперировать гаплогруппами – родами человеческими, и гаплотипами. То есть возникает возможность оперировать объективными генеалогическими «паспортами», которые тысячелетиями и десятками тысяч лет передаются по наследственной цепочке и позволяют видеть древнюю историю, древние миграции в пространстве и во времени. Такой подход дает принципиально новую возможность открыть, описать, и убедить.
На чем же основывается ДНК-генеалогия?
ДНК-генеалогия – новая наука, основы которой созданы в последние годы. Идет прогрессивное накопление экспериментального материала. Новый материал поступает потоком, ежедневно в базы данных включаются десятки и сотни новых «экспериментальных точек», которые по принципу обратной связи корректируют методологию новой науки, что приводит к уточнению методов расчета.
Экспериментальные данные ДНК-генеалогии – это картина мутаций в нерекомбинантных участках мужской половой хромосомы (на самом деле – и в митохондриальной ДНК, но в этом рассказе речь пойдет только о Y-хромосоме, расчетный и интерпретационный аппарат которой значительно более разработан). Эту картину мутаций ДНК-генеалогия рассматривает как в Y-хромосомах отдельных людей, так и их групп, популяций. Методология новой науки – перевод динамической картины мутаций в хронологические показатели, во времена жизни общих предков популяций, а на самом деле – общих предков древних родов и племен. То есть, фактически производится расчет времен, когда в древности жили эти роды и племена. В свою очередь, построение «карты» этих времен по регионам, материкам, континентам позволяет понять миграционные пути наших предков. В отрыве от археологических, антропологических, лингвистических данных ДНК-генеалогия не столь эффективна, поскольку картина мутаций и хронологические расчеты не привязаны к определенным территориям, и часто допускают множественные толкования. Но в совокупности с данными других исторических и сопряженных наук материалы ДНК-генеалогии приводят к синергизму в выводах, усиливают выводы, подвергают их перекрестной проверке под неожиданными и необычными углами, и в итоге позволяют отвести одни выводы, сделанные преждевременно и некритично, пусть и «устоявшиеся» в науке, и усилить другие.
Мутации, рассматриваемые в ДНК-генеалогии – это или одиночные (как правило) замены нуклеотидов в ДНК, как, например, аденин на цитозин, или цитозин на тимин, или вставки нуклеотидов, или делеции, или мутации более сложные, при которых ошибка копирующего фермента приводит к переносу целого блока нуклеотидов. Это дает или удлинение серии таких блоков на один (редко – сразу на два-три блока), или их укорачивание. Эти мутации наследуются в поколениях, порой в сотнях и тысячах поколений подряд без изменений, или с небольшими вариациями, и тем самым служат надежными, порой «вечными» метками в ДНК.
Мутации первого типа – SNP (Single Nucleotide Polymorphism), или «снипы», обычно очень стабильны, и их для ДНК-генеалогии отбирают только такие, которые случаются только один раз (два раза – максимум) за историю человечества. Поэтому они являются маркерами родов человечества. Эти роды называют «гаплогруппами», каждая из которых состоит из многих выявленных подгрупп, но все подгруппы несут родо-определяющую (или гаплогруппу-определяющую) SNP-мутацию. Всего в мире насчитывают 20 основных родов, обозначаемых буквами латинского алфавита от А до Т.
Мутации второго типа – STR (Short Tandem Repeat) – значительно более быстрые, и происходят в определенных локусах ДНК (то есть в гаплотипах, которые и есть совокупность выбранных локусов) раз в несколько десятков или сотен поколений. Поэтому гаплотипы ДНК выбирают так, чтобы в них таких локусов было включено как можно больше (но все же оставаясь в рамках практичности). В ранних работах использовались 6-маркерные гаплотипы, затем 12-маркерные, затем 25- и 37-маркерные, а сейчас работа рутинно ведется с 67-маркерными гаплотипами (в академических публикациях, как правило, от 8 до 17 маркерных гаплотипов), а в последние два года – и с 111-маркерными гаплотипами. В 67-маркерных гаплотипах, например, одна мутация происходит в среднем за 8 поколений, в 111-маркерных – за 5 поколений, то есть примерно раз в 125 лет. Для расчетов за «поколение» принимается интервал времени в 25 лет, и на основании этого интервала происходит калибровка констант скоростей мутации «второго типа», то есть мутаций в гаплотипах.
В этом рассказе были определены 20-маркерные гаплотипы, имеющие несколько нестандартный формат, поэтому их расчет проводился по 17 локусам, так называемому Y-filer, суммарная константа скорости мутации которых хорошо известна.
Поскольку уже доказано, что эти мутации в локусах происходят в основном (или исключительно) неупорядоченно, то к ним оказалось возможным применять правила и подходы химической кинетики (или биологической кинетики, которая базируется на тех же принципах химической кинетики, только в биологических объектах). Каждому маркеру свойственна определенная константа скорости мутации (измеряемая в числе мутаций на маркер на 25 лет, то есть на условное поколение), и каждому гаплотипу (состоящему из набора маркеров) – тоже свойственна определенная средняя константа скорости мутации. Поэтому чем древнее общий предок популяции, тем больше мутаций накапливается в гаплотипах его потомков по сравнению с ним, тем больше поколений (и лет) отделяет современных потомков от их общего предка, и это число поколений (и лет) рассчитывается методами ДНК-генеалогии. Проблема заключается в том, что начиная примерно с 600 лет до настоящего времени зависимость между общим числом мутаций и временем жизни общего предка становится нелинейной за счет накопления возвратных мутаций, и нужны специальные методологические приемы, чтобы переводить эту нелинейность в хронологические показатели. Арсенал этих приемов отличает ДНК-генеалогию, как по сути дела историческую науку (собственно, генеалогия и есть история), опирающуюся на количественный анализ кинетики накопления мутаций в ДНК, от популяционной генетики, основной задачей который является нахождение связи между генотипом и фенотипом популяции, и генетическим описание главным образом современных популяций в различных регионах.
В классификации ДНК-генеалогии рода, они же гаплогруппы, различаются по мутациям: все без исключения «восточные славяне» (R1a у славян) имеют снип М198 – мутацию цитозина в тимин в определенном участке Y-хромосомы; все «северо-западные славяне» (I1 у славян) имеют снип М253 – мутацию цитозина в тимин в другом участке; все «южные славяне» (I2a1b у славян) имеют снип L460 – мутацию аденина в цитозин в соответствующем участке; все «южно-балтийские славяне» и «угро-финны» – снип М46 (Tat) – мутацию тимина в цитозин в конкретном участке (хотя многие южно-балтийские славяне имеют еще одну мутацию, L550 – мутацию цитозина в тимин – что дает субклад N1c1a1a1, которой нет у финно-угорской ветви).
Перечисленные рода разошлись – от своих общих предков – десятки тысяч лет назад, и вот опять для миллионов людей сошлись у славян. Каждый из перечисленных родов имеет свою структуру, свои ветви популяций, каждая из ветвей имеет свою историю. Например, в Европе насчитывается 38 ветвей гаплогруппы R1a, различающихся по характерной картине мутаций в гаплотипах. Та же гаплогруппа R1a имеет 26 подгрупп, каждая со своим индексом, по состоянию на конец 2012 г.). Самая разветвленная подгруппа (субклад) – Z280, «Центральный евразийский субклад», в который входят шесть SNP подгрупп, каждая – со своим характерным мутацией-снипом, и десять ветвей гаплотипов. В принципе, им должны соответствовать десять SNP мутаций, поскольку характерные ветви должны иметь характерные снипы, но работа по поиску снипов продолжается, и часто поступают новые идентифицированные необратимые мутации, служащие, как было указано, метками, маркерами определенных популяций, фактически расширенных «семей».
Другая характерная гаплогруппа у этнических русских – I, причем в двух подгруппах, I1 и I2, представленных среди славян неодинаково (см. предыдущие главы). Обе подгруппы распространены по всей Европе, причем I1 больше на севере Европы, I2 – больше на юге, в особенности на Балканах, где она часто является наиболее представительной по численности гаплогруппой, потому ее наиболее часто встречающийся там субклад (почти 100 % от балканских гаплотипов группы I2) получил название «динарского» (индекс I2a1b). Этот же субклад почти исключительно представлен у всех славян Восточной Европы. Гаплогруппа I2, наряду с I1 – древнейшие гаплогруппы Европы, но примерно 4500 лет назад носители как I1, так и I2 из Европы исчезли, их остатки бежали на Британские острова и на Русскую равнину, и на двух концах Европы эти гаплогруппы постепенно возрождались. В популяционных науках это называется «прохождением бутылочного горлышка» популяции. Сама же гаплогруппа I появилась (путем соответствующей мутации М170 – мутация аденина в цитозин – у представителя родительской гаплогруппы IJ в соответствующем участке Y-хромосомы) примерно 40 тыс. лет назад, и примерно 30 тыс. лет назад она разошлась на подгруппы I1 и I2.
У обеих гаплогрупп тоже много SNP-подгрупп, у I1 по современной классификации 24 субклада, у I2 – 33 (по состоянию на конец 2012 г.). Но на Русской равнине выявлен почти исключительно «динарский» субклад I2a1b-M423, который впоследствии разошелся на три нижестоящих субклада – M359.2, L161.1, и L621, причем и последний уже показал «дочерний» субклад L147.2.
Наконец, еще одна из наиболее представленных у этнических русских гаплогрупп, N1c1, встречается в среднем в 14 % случаев, но от Пскова и выше ее уже более 30 %. Соответственно, на юге России этой гаплогруппы мало. Она появилась на Русской равнине последней, в начале-середине 1 тыс. до н. э., пройдя длинный миграционный путь с Урала, а туда – со стороны Алтая через Сибирь. Заняло это по меньшей мере 8 тыс. лет, а сама гаплогруппа N разошлась с гаплогруппой О из сводной (родительской) гаплогруппы NO-M214 примерно 20 тыс. лет назад, когда у одного из носителей NO в Y-хромосоме произошла мутация М231 (rs9341278), которая и положила начало 20-тысячелетней истории рода N.
Теперь подходим к белорусам, важной и неотъемлемой части славян. Из сведений, уже накопленных ДНК-генеалогией, можно ожидать, что структура белорусской популяции в целом вряд ли будет заметно отличаться от русской, украинской или польской. Равно и в отношении истории появления основных гаплогрупп на территории Беларуси нет оснований ожидать, что они будут заметно более древними или заметно более молодыми, чем на сопряженных территориях. Общие закономерности заселения европейской территории уже проанализированы и опубликованы, но генетических сведений об этнических белорусах среди них крайне мало.
И вот нам повезло. Оказалось, что ученые Национальной академии наук Беларуси и Центра судебных экспертиз Министерства юстиции Беларуси с участием Института этнографии и фольклора и Института генетики и цитологии НАН Беларуси, собрали большое количество, больше тысячи гаплотипов этнических белорусов. Это в наше время – грандиозное достижение. Работа по исследованию этнических белорусов очень важна, она отражает не только уровень науки в стране, но и понимание мировых приоритетов в научных исследованиях. Работа такого уровня дает возможность для достаточно глубоких обобщений в отношении познания не столько современной структуры белорусского народа (этим занимается популяционная генетика), сколько его этногенеза и связей с другими славянскими народами под углом ДНК-генеалогии.
Таким образом, этот очерк был бы невозможен без плодотворного сотрудничества с белорусскими коллегами, и в первую очередь с И.С. Цыбовским (первый заместитель директора Центра судебных экспертиз и криминалистики Министерства юстиции Республики Беларусь, г. Минск), и его сотрудников А.В. Богачевой, С.А. Котовой, Т.В. Забавской, Н.С. Шахнюк, а также с моими коллегами по Академии ДНК-генеалогии И.Л. Рожанским и А.С. Золотаревым. Большая часть расчетов и построения графиков в этом очерке проведена И.Л. Рожанским, за что ему огромная благодарность.
Образцы ДНК собирали по всей Беларуси, в ее центральном регионе, и по всем направлениям – на севере, юге, западе и востоке (см. далее табл.).
Когда мы построили первое дерево гаплотипов Беларуси по 312 гаплотипам, то сразу прояснилась общая структура белорусской популяции. Ветви тут же характерно разошлись по гаплогруппам, проявились четыре ветви гаплогруппы R1a, две ветви гаплогруппы R1b (древняя, с Русской равнины, и более недавняя, с запада, из Европы). Стало ясно, что толк из анализа будет, тем более из анализа более тысячи гаплотипов.
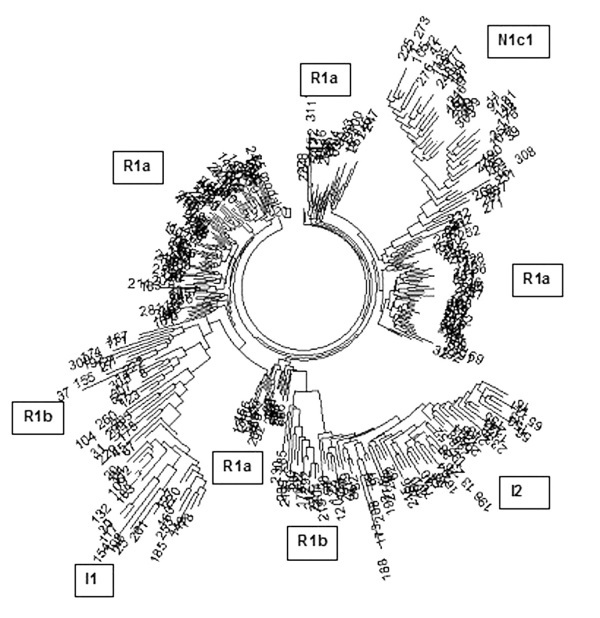
Дерево из 312 20-маркерных гаплотипов по шести регионам Беларуси. Показаны основные гаплогруппы
Приступим.
Гаплогруппа R1a (L120)
Из общего количества в почти 1100 гаплотипов в составе гаплогруппы R1a оказались 551 гаплотип, то есть 51 %. Это совпадает в целом со статистикой по соседним славянским народам – русским, украинцам и полякам. Расчет по 545 гаплотипам (которые не имели пропусков) дал в сумме 3143 мутации от следующего базового гаплотипа:
13 25 16 10 11 14 12 12 10 13 11 30 15 14 20 11 11 15 11 23.
Это соответствует 5100±520 годам до общего предка, что в пределах погрешности совпадает с величиной 4550±475 лет назад, полученные тем же методом для 258 17-маркерных гаплотипов русских, о чем шла речь в главе 7 настоящей книги.
Но более внимательное рассмотрение показало, что возраст общих предков один и тот же, просто у белорусов и русских немного различаются субклады: у белорусов больше западнославянского субклада (М458), у русских – центрально-евразийского субклада), и вот расчеты «поперек ветвей» и привели к кажущейся разнице в возрасте популяций. Впрочем, все равно в пределах погрешности.
Было бы хорошо провести расчеты по всем ветвям гаплогруппы R1a, но с 20-маркерными гаплотипами это почти невозможно, нужны 67-маркерные. Но, тем не менее, четыре ветви у белорусов удалось идентифицировать. Они показаны на дереве гаплотипов гаплогруппы R1a. Это – центрально-европейская ветвь (М458-L1029), 2900±400 лет от общего предка, базовый гаплотип
13 25 16 10 11 14 12 12 11 13 11 29 16 14 20 11 11 17 11 23,
западнославянская (М458-L260), общий предок жил 2700±300 лет назад
13 25 17 10 10 14 12 12 10 13 11 30 16 14 20 11 11 16 11 23,
северно-евразийская (Z280-Z92), 2350±300 лет до общего предка,
13 25 16 11 11 15 12 12 10 13 11 30 15 14 20 11 12 15 11 24
и балто-карпатская (Z280), 4300±500 лет до общего предка
13 25 16 10 11 14 12 12 10 13 11 30 15 14 20 11 11 16 11 23.
Последняя ветвь характерна для Прибалтики, а также польского Поморья, и ее наличие у белорусов вполне закономерно.
Другие принципиальные субклады – «восточный» R1a1a1b2 (Z93), скандинавский R1a1a1b1a3 (Z284) и северо-западный R1a1a1a (L664), присутствуют у всех славянских народов на уровне долей процента, и крайне маловероятно, что белорусы составляют исключение.
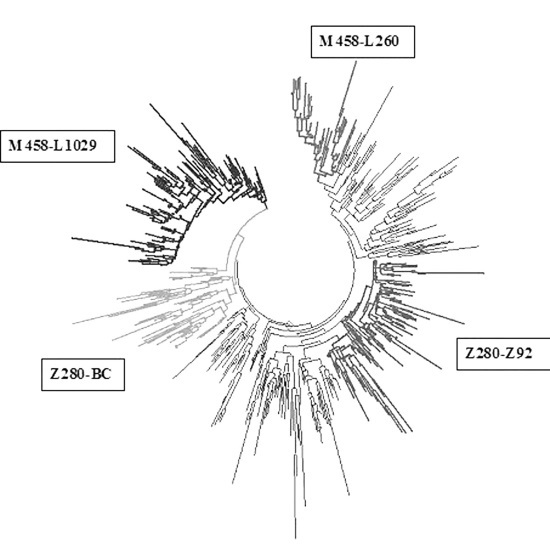
Дерево 551 гаплотипа гаплогруппы R1a. По часовой стрелке, начиная сверху, показаны ветви западнославянская (М458-L260, 36 гаплотипов), северная евразийская (Z280-Z92, 128 гаплотипов), балто-карпатская (Z280, специфическая снип-мутация пока не определена, 88 гаплотипов) и центрально-европейская (М458-L1029, 131 гаплотип)
Расчеты дали время до общего предка белорусских гаплотипов группы R1a (как время возникновения общего предка субкладов Z280 и M458), равное 4800±500 лет, а для русских (в основном) это время, рассчитанное по 67-маркерным гаплотипам, равно 5500±600 лет назад, что то же самое (в пределах погрешности). Тем самым положение об одном и том же общем предке носителей R1a у русских и белорусов можно считать доказанным. Естественно, пять тысяч лет назад он не был ни русским, ни белорусом. Вскоре его род двинется на восток, заселит территории современных Беларуси и России, а также сопряженных территорий, и его потомки станут в итоге современными белорусами и русскими гаплогруппы R1a.
Гаплогруппа I (М170)
Сводная гаплогруппа I занимает у белорусов второе место после R1a, охватывая 24 % от всей выборки. Дерево имеет довольно простую структуру, и распадается на две однородные ветви с относительно недавними общими предками, а также небольшую (<1 % от выборки) группу гаплотипов из других ветвей I2.
Выборка гаплогруппы I1 у белорусов сходится к общему предку, жившему 3700±450 лет назад, и к базовому гаплотипу:
13 23 14 10 14 14 11 14 11 12 11 28 15 16 20 10 10 14 10 22.
Это совпадает с данными по всей гаплогруппе при расчете по протяженным гаплотипам. В 20-маркерном формате ветвь не удается разрешить на дочерние, а потому в данном приближении белорусские I1 можно считать неотличимыми от остальных, которые распределены в основном в северной части Европы.
Еще более однородной является ветвь I2a1b, представленная 194 гаплотипами. Базовый гаплотип ветви:
13 24 16 11 14 15 11 15 13 13 11 31 17 15 20 10 10 15 10 23.

Дерево 262 гаплотипов гаплогруппы I. Показаны гаплогруппа I1 (М253, 61 гаплотип), «динарская» ветвь I2a1b (M423, 194 гаплотипа), и ветви I2, отличные от «динарской» (10 гаплотипов)
Он совпадает с базовым гаплотипом «динарской» ветви, повсеместно встречающейся у славян и достигающей пика распространенности в Боснии и Герцеговине. время до общего предка равно 2200±250 лет. Следовательно, общий предок всех носителей «динарской» ветви I2a1b, вне зависимости от региона, один и тот же, с временем жизни 2200±250 лет назад (см. предыдущую главу книги).
Гаплогруппа N1c1 (Tat).
Подобно ветвям гаплогруппы I, белорусская выборка из 109 гаплотипов N1c1 дает довольно однородное дерево, в котором сложно выделить дочерние подветви. Ее базовый гаплотип:
14 23 15 11 11 13 11 12 10 14 14 30 17 14 19 11 11 14 10 21.
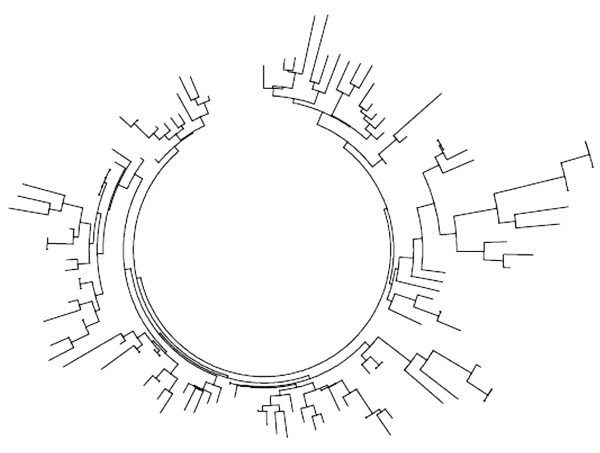
Дерево 109 гаплотипов гаплогруппы N1c1
Это – южно-балтийская ветвь (N1c1a1a1-L550), которая в полном 67-маркерном формате имеет следующий базовый гаплотип (отмечены совпадающие аллели):
14 23 14 11 11 13 11 12 10 14 14 30–17 9 9 11 12 25 14 19 28
14 14 15 15–11 11 18 20 14 15 16 19 35 35 14 10–11 8 15 17
8 8 10 8 11 10 12 21 22 14 10 12 12 17 7 13 20 21 16 12 11 10 11
11 12 11 (южно-балтийский базовый гаплотип).
В численном отношении доля гаплогруппы N1c1 среди белорусов составляет 10 %.
Это заметно меньше, чем среди этнических русских (14 %), но больше, если принимать во внимание только ее южно-балтийскую ветвь (40–50 % от N1c1 у русских, то есть 6–7 %).
Гаплогруппа R1b1a2 (M269)
К самой распространенной в Западной Европе гаплогруппе R1b1a2 относится 58 гаплотипов из белорусской выборки (5,3 %), что практически совпадает со статистикой у русских (4,8 %). Дерево распадается на две ветви, по 29 гаплотипов каждая. Первая сходится к общему предку, жившему 3725±520 лет назад. Ее базовый гаплотип:
12 24 14 11 11 14 12 12 12 13 13 29 16 15 19 11 11 15 12 23.
Вторая ветвь имеет практически тот же «возраст», 3825±520 лет, но ее базовый гаплотип отличается на 5 мутаций:
13 23 14 10 11 14 12 12 12 13 13 29 17 15 19 11 11 16 12 23.
Первый базовый гаплотип совпадает с соответствующим фрагментом 67-маркерного базового гаплотипа ветви R1b1a2a1 (L150+, L51-) с предком, жившим около 6000 лет назад. Она крайне редко встречается в Западной Европе, но характерна для ряда народов Северного Кавказа и Закавказья (армяне, грузины, ассирийцы, осетины, дагестанцы), Восточной Европы (башкиры, татары, чехи, греки) и Центральной Азии (туркмены, казахи, уйгуры). Второй базовый гаплотип – это один из вариантов европейских ветвей субклада R1b1a2a1a (L51+), которых в текущей классификации насчитывается 155. Разница в 5 мутаций между «западным» и «восточным» базовыми гаплотипами соответствует 4300 годам между ними, и их общему предку, жившему (3725+3825+4300)/2?5900 лет назад, что в пределах погрешности совпадает со временем жизни предка родительского субклада R1b1a2a1 (L150). Относительно недавнее время жизни предка «восточной» ветви у белорусов, возможно, говорит, что среди них преобладают представители какой-либо из дочерних ветвей этого субклада, пока недостаточно изученного в сравнении с западноевропейскими ветвями.
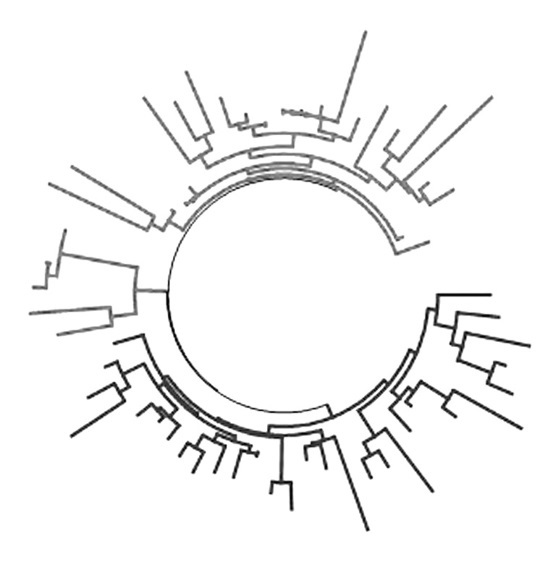
Дерево 58 гаплотипов гаплогруппы R1b1a2. В верхней части находится ветвь, предположительно относящаяся к «восточному» субкладу R1b1a2a1 (L150), в нижней – гаплотипы из западноевропейского субклада R1b1a2a1a (L51)
Гаплогруппа E1b1b1 (M35.1)
Процент носителей гаплогруппы E1b1b1 у белорусов относительно невелик (4 %), на том же примерно уровне, что у других европейских народов, не относящихся к средиземноморскому ареалу. Базовый гаплотип:
13 24 13 10 16 18 11 12 12 13 11 30 15 14 20 9 11 16 10 21
совпадает с базовым гаплотипом ветви E1b1b1a1b (V13), наиболее представленной в Европе.
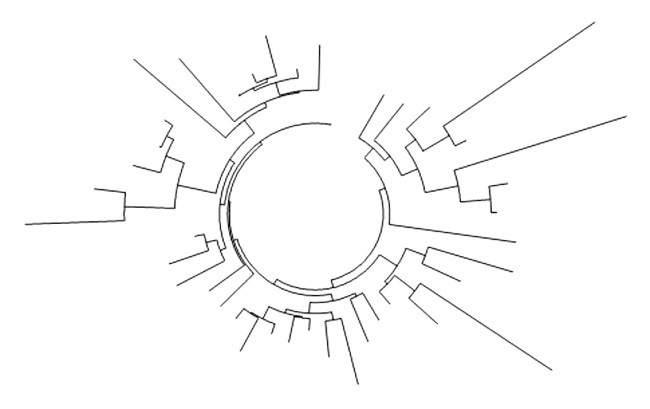
Дерево 41 гаплотипа гаплогруппы E1b1b1
Гаплогруппа J (Р209)
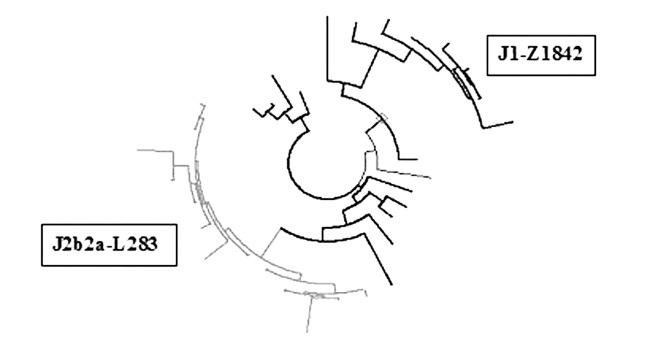
Дерево 33 гаплотипов гаплогруппы J. Справа вверху находится не имеющая пока индекса ветвь гаплогруппа J1 (Z1842) (8 гаплотипов); слева внизу – ветвь J2b2a (L283) (13 гаплотипов)
К гаплогруппе J в выборке относятся 33 гаплотипа (3,0 % от всех). Выделяется ветвь слева внизу из 13 гаплотипов с предком, жившим 3100±600 лет назад, и базовым гаплотипом:
12 24 15 10 13 17 11 15 12 12 11 28 16 16 19 11 10 13 9 21.
Он всего на одну мутацию (выделена) отличается от базового гаплотипа ветви J2b2a (L283), имеющей «возраст» 4000±450 лет и рассеянной с низкой частотой по всей Европе, но почти не встречающейся на Ближнем Востоке. Очевидно, 13 белорусов относятся к той же самой довольно редкой европейской ветви гаплогруппы J2.
Следы еще одной редкой генеалогической линии обнаруживаются среди носителей гаплогруппы J1, которые образуют довольно компактную ветвь с базовым гаплотипом:
13 24 14 10 13 19 11 13 12 13 11 29 19.2 14 21 11 11 15 10 21.
Общий предок попадает на время 3000±650 лет назад, а базовый гаплотип этой малочисленной ветви на 5 мутаций (выделены) отстоит от базового гаплотипа ветви J1*(DYS388=13), имеющей «возраст» около 5000 лет и характерной для народов Северного Кавказа. Разница в 5 мутаций в данном формате соответствует 4300 годам между гаплотипами, и общий предок белорусской и северокавказской выборок датируется временем (3000+5000+4300)/2=6150 лет назад. Следовательно, носители гаплогруппы J1 в выборке из Беларуси представляют ветвь, родственную или дочернюю к «кавказскому» субкладу.
Вопреки ставшему штампом определению гаплогруппы J1 как семитской, рано отделившаяся ветвь Z1842 практически не имеет в своем составе ни арабов, ни евреев, а потому крайне маловероятно, что рассматриваемые 8 гаплотипов – след евреев ашкенази, составлявших до Первой мировой войны до 20 % населения Беларуси. У нее другое происхождение, пока неизвестное.
Остальные гаплотипы, что относятся, по-видимому, к разным ветвям субклада J2a, группируются по 2–3, не образуя выраженных ветвей.
Гаплогруппы C3, G1, G2a, N, Q, R1b1a1, R2, T
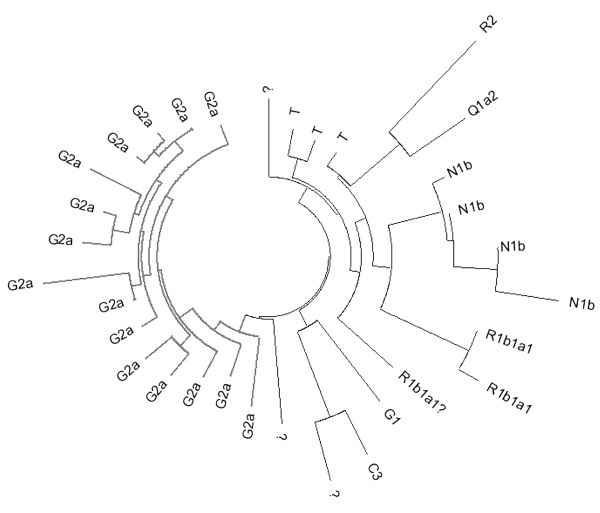
Дерево 32 гаплотипов из минорных гаплогрупп, перечисленных в названии раздела
На долю всех остальных гаплогрупп приходится 32 гаплотипа или 3 % от всей выборки. Из них 15 относится к гаплогруппе G2a. Их базовый гаплотип можно записать как:
14 22 15 10 14 15 11 12 11 12 11 29 17 16 21 11 11 15 10 21.
Он совпадает с базовым гаплотипом родительской ветви субклада G2a1c2a (Р303), характерного для Северного Кавказа, но рассеянного с низкой частотой по всей Европе и, судя по находкам ископаемой ДНК, представлявшего одну из основных генеалогических линий Европы эпохи неолита. Время до общего предка белорусских гаплотипов (7200±1100 лет) совпадает в пределах погрешности с оценкой для субклада Р303 (около 6300 лет назад). Очевидно, если не все, то значительная часть носителей этой гаплогруппы в Беларуси относится к субкладу G2a1c2a.
Единичные гаплотипы из других ветвей собраны в правой части дерева. Из них, как минимум, 9 гаплотипов можно отнести к линиям центрально- и восточноазиатского происхождения. Это гаплогруппы С3 (М217), G1 (M342), N1c2b (Р43), Q1a2 (М25) и R1b1a1 (М73). Их крайне мало, на уровне долей процента, как и у русских. Да и те, что есть, возможно, достались частично от литовских татар, переселение которых из Крыма в Великое княжество Литовское в XV веке хорошо документировано.
Татары рано утеряли свой язык, перейдя на старобелорусский, а часть их крестилась, слившись тем самым с белорусами.
* * *
Итак, у белорусов представлены те же самые Y-хромосомные линии, что и у русских, с теми же самыми общими древними предками. Следовательно, оба восточнославянских народа близки по своему происхождению, поскольку восходят к одним и тем же группам древних этносов. Различия касаются в основном процентного соотношения этих линий, а также географического распределения для некоторых из них. Следует понимать, что хотя выявленные в генетической структуре этнических белорусов и русских гаплотипы сводятся во времени к общим древним предкам (с приведенными базовыми гаплотипами), однако это не означает, что данные предки проживали (и проживали ли?) на территориях, занимаемыми современными этносами. Вопрос, какие конкретно из потомков древних генеалогических линий в конце концов осели на той или иной территории, более сложный. Анализ географического распределения Y-хромосомных линий более четко иллюстрирует данную проблему.
Статистически значимые результаты такого анализа можно получить из табл. 1, где собраны данные по славянским народам, а также ближайшим соседям белорусов – литовцам.
Таблица 1. Распределение основных гаплогрупп и ветвей у славян и литовцев (%) [по данным И.Л. Рожанского и белорусского научного коллектива криминалистов]. A – юго-запад Беларуси, В – северо-запад, С – центр и север, D – юг и юго-восток
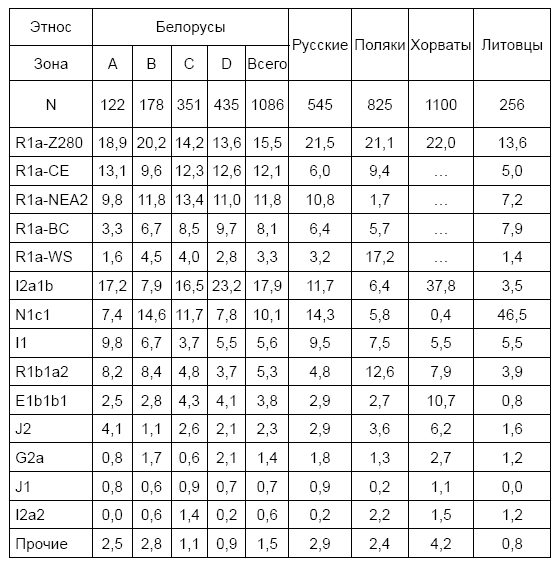
Обозначения ветвей R1a: CE – центрально-европейская, WS – западнославянская, NEA2 – северная евразийская (Z280+, Z92+), подветвь с предком 2350±300 лет назад, BC – балто-карпатская, Z280 – остальные ветви субклада, неразличимые в 20-маркерном формате. Статистика по полякам получена с польского проекта FTDNA. Распределение гаплотипов R1a по ветвям было получено путем анализа базы данных IRAKAZ, содержащей 3126 гаплотипов R1a длиной 67 маркеров и более
Поскольку распределение некоторых генеалогических линий на территории Беларуси имеет явно выраженный градиент (I2a1b, R1a-Z280, табл. 1), использование при анализе усредненных значений будет заведомо некорректным. По этой причине места сбора образцов были поделены на три зоны: зона B – северо-западная территория, непосредственно примыкающая к Литве; зона C, занимающая центр Беларуси и часть Поозерья; и зона D, охватывающую Полесье и южную часть Поднепровья. В дополнение к ним, три самых западных места сбора – Волковыск, Кобрин и Береза (часть Западного Полесья и южная часть Понеманья), были выделены в зону A.
В распределении ветвей по зонам хорошо видно особое положение зоны B, граничащей с Литвой. Там заметно снижен процент носителей ветви I2a1b (8 % против 18 % в среднем по Беларуси) и повышено представительство N1c1 (15 и 10 %, соответственно). По мере удаления от Литвы (зоны C и D) процент носителей N1c1 закономерно снижается, а I2a1b – растет. Если сравнить эти данные со статистикой по литовцам (4 % I2a1b и 47 % N1c1), то очевидно, что такую тенденцию можно объяснить вкладом древних балтских этносов в этногенез белорусов. Об этом говорит и тот факт, что в коммерческих базах данных носители гаплогруппы N1c1 как у литовцев, так и у белорусов, представлены исключительно южно-балтийской ветвью (L550+), в то время как у русских, в состав которых входят потомки ассимилированных финно-угорских народов, эта ветвь составляет не более 40–50 % от всех носителей N1c1. Эти данные также позволяют предположить, что вклад финно-угорских народов Русской равнины в этногенез белорусов был минимальным, если не отсутствовал вообще.
Ветви гаплогруппы R1a не показывают статистически значимых корреляций с географией – они распределены по территории Беларуси достаточно равномерно. Несколько особое положение занимает северная евразийская ветвь (Z92) – одна из основных генеалогических линий белорусов, русских и литовцев, но редко встречающаяся у западных и южных славян. В зоне A, граничащей с Польшей, можно также отметить более высокую долю носителей гаплогрупп I1 и R1b1a2 – 10 и 8 % против 6 и 5 % в среднем по Беларуси, соответственно. Остальные генеалогические линии, что сообща охватывают 10 % белорусов, слишком малочисленны, чтобы делать подобные оценки.
Белорусы занимают «стратегически» важный регион между балтами на севере и украинцами на юге, между поляками и русскими на западе и востоке, соответственно, и к северу от Карпат, по которым в древности проходили многочисленные миграции славянских племен. Совместный анализ датировок и распределения генеалогических линий у белорусов и соседних народов дает возможность очертить несколько волн заселения, которые внесли свой вклад в этногенез современных белорусов. Самые ранние датировки прослеживаются примерно с 4300 лет назад, когда начался рост балто-карпатской ветви R1a, а также родительских ветвей западной и центральной евразийской ветвей того же субклада Z280, К той же волне, скорее всего, относится и большинство носителей гаплогруппы I1 у белорусов и русских. Эту волну можно условно назвать автохтонной, поскольку генеалогические линии тех, кто ранее жил на этой территории до того, пресеклись, и их Y-хромосомные гаплогруппы нам неизвестны.
Следующая по счету волна связана с ростом южно-балтийской ветви N1c1, начавшимся 2700±300 лет назад. В настоящее время пик распространенности этой ветви приходится на Литву и Латвию, но это не обязательно означает, что ее предок жил именно там. Гаплогруппа N1c1 имеет азиатское происхождение, а потому не менее вероятно, что ее южно-балтийская ветвь сформировалась где-то восточнее, и на берега Балтийского моря ее носители пришли несколько позже. Поскольку пути и датировки миграции соответствующих этносов пока неизвестны, эту волну затруднительно соотнести с какой либо из современных ей археологических культур.
Наконец, третья ключевая датировка попадает на середину-конец 1 тыс. до н. э. Она соответствует быстрому росту «динарской» ветви I2a1b (2200±250 лет назад), северной евразийской ветви R1a (2350±250 лет назад), а также нескольких дочерних ветвей субклада Z280, что не выделяются на дереве 20-маркерных гаплотипов, но были охарактеризованы по протяженным гаплотипам. Судя по географическому распределению этих линий, их рост шел из разных центров. Скорее, это были встречные миграции с юга или юго-запада (I2a1b и часть ветвей Z280) и со стороны Балтики (северная евразийская ветвь R1a). Очень возможно, что последняя зародилось в Беларуси.
Не совсем ясна картина с субкладом М458, ветви которого начали выделяться раньше (3000–2700 лет назад), но, судя по пику в Чехии и Польше, а затем спаду их численности в восточном направлении, носители этого заметно отстоящего от Z280 субклада могли появиться на территории Беларуси позже. Возможно, с той же волной расселения, что и упоминавшиеся выше ветви. Представители этих генеалогических линий составляют сейчас основу всех без исключения славянских народов, как современных, так и подвергшихся ассимиляции, и влившихся в состав немцев, венгров, румын, греков, турок. Есть все основания связать эту цепь демографических событий с миграциями славян и распространением славянских языков.
Как видно из анализа датировок и географии, сомнительно, чтобы все эти родственные, но все же разные народы подпадали под одну и ту же археологическую культуру. Этим, видимо, и объясняются безуспешные попытки привязать прародину славян к какому-то конкретному месту и дате. Их формирование было растянуто во времени и пространстве, как можно судить по сумме доказательств. Детали этого процесса еще предстоит выяснить, но несомненно, что территория Беларуси играла в нем одну из ключевых ролей. В любом случае, Беларусь составляет единую славянскую зону со славянами Восточной Европы по данным ДНК-генеалогии с точки зрения истории миграций и времен жизни общих предков ДНК-генеалогических линий.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК