Распространенье наше по планете
Распространенье наше по планете
Примерно 1,7 млн. лет назад, т. е. в самом начале эпохи, которую геологи называют плейстоценом или ледниковым периодом, в истории семейства гоминид и рода Homo случилось ещё одно знаменательное событие — его представители впервые проникли за пределы Африки. Сначала они обосновались на Ближнем Востоке и в Закавказье, а затем постепенно стали расселяться всё дальше на восток, не выходя, однако, слишком далеко за пределы привычной для них зоны тропиков и субтропиков. Миллион лет назад ими был заселен уже почти весь юг Азии, включая территорию нынешнего острова Ява. Примерно к этому же времени относятся, вероятно, и первые попытки обосноваться в Европе, где возраст древнейших достоверно человеческих костей, а также каменных орудий достигает 1,2 млн. лет.
Пионерами-первопроходцами в освоении дальних земель были не габилисы, а те, кого раньше несколько уничижительно именовали питекантропами (т. е. обезьянолюдьми), а сейчас относят к виду гомо эректус (Homo erectus). Иногда этот вид делят на два, и более раннюю африканскую форму называют гомо эргастер (Homo ergaster). Для этого вида, появившегося где-то 1,8–1,9 млн. лет назад, в общем, характерен уже вполне человеческий скелет, который по многим параметрам, в том числе по размерам и пропорциям, очень близок к современному состоянию. Особенно показательно в этом смысле соотношение длины бедренной, плечевой и предплечевых костей. У шимпанзе оно равняется примерно 1:1, а у людей, хотя плечо имеет почти тот же абсолютный размер, бедро удлинено, а предплечье укорочено. Впервые эти человеческие пропорции фиксируются именно у гомо эректус. Череп на этой стадии эволюции человека тоже изменяется, и возрастает объём мозговой полости. Средний размер эндокрана эректусов составляет около 950 см3, при крайних значениях от 700 до 1200 см3.
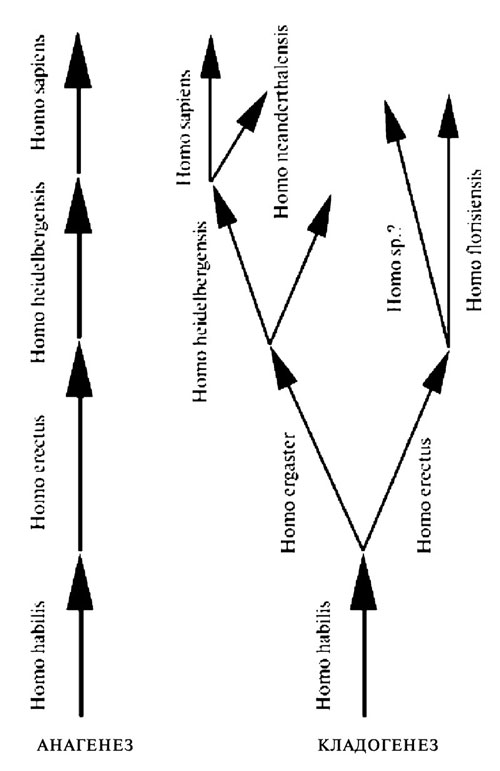
Рис. 3.3. Возможные варианты анагенетической и кладогенетической эволюции рода Homo. Сторонники крайней версии первого подхода иногда сокращают число видов до двух (Homo habilis и Homo sapiens)
После широкого расселения за пределы Африки между разбросанными в пространстве человеческими популяциями стало постепенно проявляться всё больше и больше различий. Непохожие природные условия Африки, Восточной Азии и Средиземноморья предъявляли к обитавшим в этих регионах гоминидам разные требования, отбор работал в разных направлениях и с разной интенсивностью. Как следствие этого, эволюционные изменения должны были вести к формированию специфических особенностей в поведении и анатомии каждого из географических вариантов гомо эректус и их потомков. Конечно, процесс расхождения в какой-то мере мог сглаживаться за счёт обмена генами, которые даже при скрещивании только между соседними популяциями со временем способны распространяться на тысячи и десятки тысяч километров, с одного края ойкумены на другой, но вряд ли этого было достаточно для поддержания биологического единства гоминид, живших на разных континентах. Вероятно, они ещё долго сохраняли потенциальную способность скрещиваться и давать при этом нестерильное потомство, т. е. оставались членами одного вида в репродуктивном смысле. Однако углубляющиеся различия в анатомии и в поведении (а также географическая изоляция) вели к тому, что на практике эта способность реализовывалась нечасто. Подобная ситуация характерна для многих современных животных, например для тигра и льва, волка и собаки, а также для целого ряда разных видов и даже родов приматов.
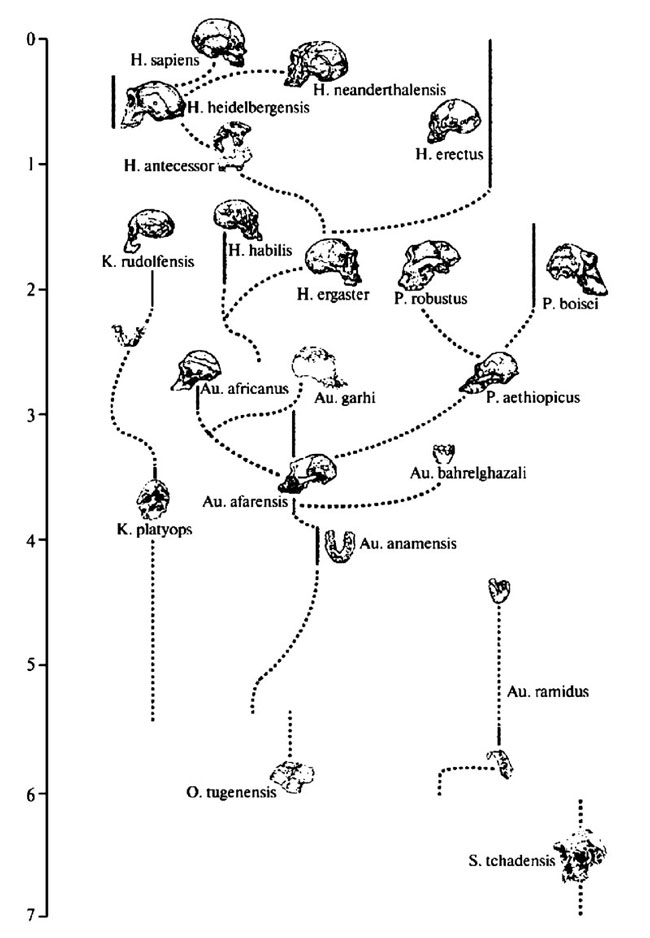
Рис. 3.4. Одна из версий генеалогического древа гоминид (источник: Tattersall 2004)
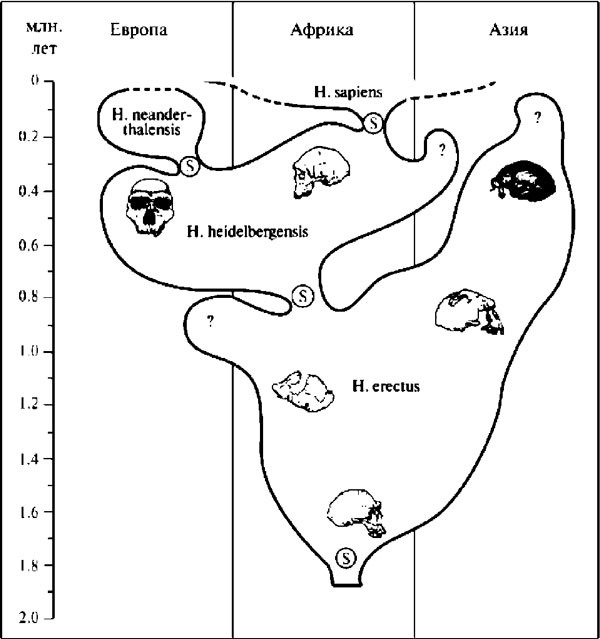
Рис. 3.5. Схема эволюции рода Homo, предполагающая, что Homo sapiens первоначально появляются в Африке, а оттуда распространяются в другие регионы, замещая предшествовавшие им там формы гоминид (источник: Rightmire 1998)
Где-то в самом конце нижнего и/или начале среднего плейстоцена, т. е. примерно 700–800 тыс. лет назад, в анатомии представителей рода Homo снова происходит ряд важных изменений. В основном они касаются строения черепа. Лоб становится более широким, теменная область тоже заметно разрастается, а затылок округляется и лишается свойственного эректусам угловатого выступа. В результате объём мозговой полости резко возрастает: отныне он обычно превышает 1100 см3, а в отдельных случаях достигает даже размеров, типичных для современных людей (1300–1400 см3). Гоминид, обладающих перечисленными признаками, как правило, уже не включают в вид гомо эректус. Большинство антропологов относит их к виду, именуемому гомо гейдельбергенсис (Homo heidelbergensis, человек гейдельбергский).
Вполне естественно, что эволюционная судьба различных географических популяций гомо эректус и пришедших им на смену гомо гейдельбергенсис сложилась по-разному. Согласно преобладающей сейчас точке зрения, часто именуемой теорией «африканского исхода», лишь африканские представители этих видов имели непосредственное отношение к происхождению людей современного анатомического типа, т. е. были прямыми предками гомо сапиенс. Азиатским и европейским гоминидам в этом случае остаётся роль боковых побегов на нашем генеалогическом древе (рис. 3.5). Таким образом, общий сценарий происхождения современного человечества получается моноцентристским. Противоположную позицию занимают исследователи, защищающие так называемую «мультирегиональную» — полицентристскую — теорию, согласно которой эволюционное превращение гомо эректус в гомо сапиенс происходило повсеместно, то есть и в Африке, и в Азии, и в Европе, при постоянном обмене генами между популяциями этих регионов. Хотя спор между моноцентристами и полицентристами, имеющий длительную историю, всё ещё продолжается, накапливающиеся новые данные гораздо естественней укладываются в рамки первой теории, утверждающей, что люди современного анатомического типа появились сначала в одном регионе (а именно — в Африке), откуда распространились впоследствии по всей земле. Подробней об этом ещё будет рассказано в главе 10.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Новый год шагает по планете
Новый год шагает по планете Хрустит под ногами податливый снег, мороз подкрашивает лица задорным румянцем, на отрывном календаре остается один только листок — 31 декабря. Сколько веселых забот, воспоминаний, надежд связано с последним днем уходящего года! А почему
НАШЕ ПОЛОЖЕНИЕ
НАШЕ ПОЛОЖЕНИЕ ПРЕДИСЛОВИЕ В классификации военных знаний искусство вести бой называется тактикой, а искусство вести войну — высшей тактикой или стратегией. Но как бой представляет собою только один из скоротечных актов длящейся обыкновенно годами войны, так и война
«Мир в наше время»
«Мир в наше время» Я так и не могу решить, верить ли эпизоду, которым Аммиан практически на последней странице своей «Истории» завершает рассказ о Готской войне. Показав победоносных готов, собирающихся осадить Константинополь, он затем преподносит нам следующее:
Глава XII Наше все
Глава XII Наше все Обыкновенная история Покой, сытость и избыток времени, известное дело, предрасполагают к философствованиям. Среди старосветских помещиков Малой Руси, тех самых, служить не желавших, а желавших грезить под бандуру с галушками, было немало людей
Наше богатство
Наше богатство Иноземные путешественники и послы, посещавшие Россию несколько столетий назад, обращали внимание на обилие дремучих лесов, которых в Западной Европе со времен Возрождения почти не осталось. На Руси в лесах добывали в огромных количествах мед диких пчел,
Глава шестая «ЭТО СТОЛКНОВЕНИЕ МОГЛО СТОИТЬ ПЛАНЕТЕ МИРА!»
Глава шестая «ЭТО СТОЛКНОВЕНИЕ МОГЛО СТОИТЬ ПЛАНЕТЕ МИРА!» Телеграмма появилась в ленте новостей на третий день после гибели «Курска»:«ВАШИНГТОН. 15 августа (Корр. ИТАР-ТАСС). Во время происшествия с российской атомной подводной лодкой «Курск» вблизи от неё находились две
Миграции праславянских языков по планете и их следы
Миграции праславянских языков по планете и их следы Из древнеписьменных источников известно, что империя Атлантида состояла из 15 конфедераций, располагавшихся по обе стороны теплого течения Гольфстрим от Южной Америки до Таймыра и Чукотки. Помимо Священной столицы в
Древнейший город на планете?
Древнейший город на планете? В Сирии обнаружены руины города, возраст которого, по мнению ученых, составляет не менее 6000 лет. Возможно, это самый старый город на планете. Находка фактически изменила традиционные представления о появлении на Земле городов и цивилизации в
О, горе наше!
О, горе наше! И выражая настроения оппозиции, Победоносцев писал в Москву все той же Екатерине Тютчевой:«…При нем (Лорис-Меликове) затихло – но увидим, надолго ли… Он поднимает и распускает силы, с которыми трудно будет справиться. Рецепт его легкий… поднялись студенты –
НАШЕ ПРОИСХОЖДЕНИЕ
НАШЕ ПРОИСХОЖДЕНИЕ Позиции и деятельность революционных организаций являются продуктом опыта, накопленного в прошлом рабочим классом, и уроков, извлеченных из истории его политическими организациями. ИКТ ведет своё происхождение от Союза коммунистов Маркса и Энгельса
Мифология и материальная культура россо-ариев и движение их культуры по планете
Мифология и материальная культура россо-ариев и движение их культуры по планете Как материально жили люди, создавшие основу мировой мифологии и поименовавшие сотни культовых рек и множество гор и урочищ? Чем россо-арии занимались кроме рыболовства?Начнем с Нави, мира