Кости животных. Зооархеология
Кости животных. Зооархеология
Ветхозаветный пророк Иезекииль невольно дал определение работе зооархеолога: «И я изрек пророчество, как Он повелел мне, и вошел в них дух, — и они ожили, и стали на ноги свои — весьма, весьма великое полчище» (Книга пророка Иезекииля 37:10). Зооархеологи буквально облекают плотью давно умерших животных, реконструируя окружающую среду и деятельность древних людей в той степени, в которой позволяют исследования по останкам животных. Зооархеология является областью знания, требующей познаний в палеонтологии и зоологии.
Зооархеология занимается изучением костей животных, обнаруженный в археологическом материале. Ее целью является реконструкция окружающей среды и деятельности древних людей в той степени, в которой позволяют это сделаты исследования по остаткам животных (Кляйн и Круз-Урибе — Klein and Cruz-Uribe, 1984). Хотя некоторые зоологи и специализируются в изучении костей животных из археологических памятников, все же большинство зооархеологов имеют образование и опыт работы в палеонтологии или в исследованиях доисторической фауны.
Тафономия
Слово тафономия (от греч. taphnos — могила; nomos — закон) используется для описания процессов, которые происходят с органическими остатками при образовании ископаемых отложений (Лаймэн — Lyman, 1994; Шипмэн — Shipman, 1981). Говоря проще, это изучение перехода остатков животных из биосферы в литосферу.
ОТКРЫТИЯ
ГИРЭМ БИНГЕМ В МАЧУ-ПИКЧУ, ПЕРУ, 1911 ГОД
«Затерянный город инков» являлся одной из археологических тайн конца XIX века, легендой о последнем оплоте инков, где их правители прятались от ненасытных испанских конкистадоров после того, как Франциско Писарро сверг их империю в 1534 году. Молодой выпускник Йельского университета по имени Гирэм Бингем попал под влияние этой тайны и проник на памятник Вилькабамба высоко в Андах, но понял, что это не то поселение. Он уговорил своих состоятельных друзей по университету финансировать вторую экспедицию в Анды.
Упрямый и чрезвычайно любопытный Бингем был опытным альпинистом и имел хорошую историческую подготовку. Он вышел из Куско в 1911 году с караваном мулов и двигался вдоль реки Урубамба, восхищаясь замечательными видами заснеженных гор, горных ручьев и тропической растительности. Случайная встреча с местным крестьянином Мелкором Артегой подарила ему рассказ о каких-то развалинах в горах за рекой. 24 июля 1911 года Бингем вместе с этим крестьянином и перуанским сержантом перешли через Урубамбу по деревянному мостику. Ошибки быть не могло. Он карабкался на четвереньках по узкой тропинке и забрался на высоту 600 метров в лесу на противоположной стороне реки. После короткого отдыха в индейском поселении он продолжил свой путь наверх. За отрогом горы он увидел недавно расчищенные каменные террасы, поднимавшиеся на 300 метров. Над террасами, которые расчистили индейцы, он попал в густой лес и очутился между зданиями, среди которых был трехсторонний храм с такой же замечательной кладкой, как в Куско или Оллантайятамбо. Он стоял перед стенами разрушенных домов, построенных с величайшим мастерством инков. Гирэм пробился через подлесок и вошел в полукруглое здание, внешняя сторона которого, чуть наклонная и слегка изогнутая, удивительно напоминала Храм Солнца в Куско. Бингем вошел в самые знаменитые из всех руин инков, Мачу-Пикчу (рис. 13.1).

Рис. 13.1. Мачу-Пикчу, Перу
Ископаемая фауна проходит через несколыко стадий, пока из биосферы попадает в руки археологов. Первоначально кости происходят из того, что ученые называют биоценозом, то есты совокупность живых животных в их естественных пропорциях. Убитые животные или умершие по естественным причинам образуют некроценоз — туши или части туш, находящихся на памятнике. Комплексы ископаемых остатков — тафоценоз — состоят из частей животных, которые сохранилисы на памятнике до раскопок. Комплекс образцов — это то, что дошло до лаборатории, та часты комплекса ископаемых остатков, что быта собрана или включена в коллекцию (Кляйн и Круз-Урибе — Klein and Cruz-Uribe, 1984). Любой человек, занимающийся анализом фауны, должен решиты две проблемы: статистическую проблему оценки характеристик комплекса ископаемых остатков по образцу и тафономическую проблему — сделаты вывод о природе некроценоза по комплексу ископаемый остатков.
В тафономии есты два родственный направления исследования. Первое — это фактическое наблюдение недавно умерших органических остатков и того, как они постепенно превращаются в ископаемое; другое направление — это изучение ископаемый остатков в свете этих сведений. Эта область исследования стала актуальной в 1960–1970-е годы, когда археологи начали интересоватыся значением россыпей костей животных на таких древних памятниках, как ущелые Олдувай в Восточной Африке, и особенно в знаменитых пещерах Australopithecus в Южной Африке (Брэйн — Brain, 1981).
На многие вопросы о процессах, которые превращают живые организмы в «археологические» кости, ответов пока нет, несмотря на некоторые исследования о том, каким образом кости могли быть перенесены и разобщены как плотоядными животными, так и естественными агентами, такими как вода. Например, опыты с гиенами, находящимися в неволе, показали, что они сначала выбирают кости позвоночника и тазовые кости, которые они обычно разрушают полностью. Концы длинных трубчатый костей конечностей часто разгрызают полностью, в то время как их тела (диафизы) нередко остаются нетронутыми. Эти опыты очень важны, потому что они свидетельствуют о том, что запас костей, сделанных древними гоминидами в ущелые Олдувай, растаскивался гиенами после того, как люди уходили. Этот процесс вел к уничтожению многих частей тела, и поэтому невозможно сказаты, выборочно ли гоминиды утаскивали части жертв хищников или нет (Мэриэн и другие — Marean and others, 1992). Люди расчленяли животных с помощью орудий до того, как туши уничтожали плотоядные или естественные процессы, поэтому систематические действия людей считаются, по крайней мере, основным фактором при изучении повреждений археологических костей. Интерпретация доисторических жилых ярусов и мест убиения животных должна проводитыся с большой тщательностью, посколыку комплексы костей и артефактов в таких местах говорят не только о деятельности человека, но также и о сложных и мало понимаемых естественных процессах.
Многие зооархеологи считают, что по комплексам костей из археологических памятников невозможно реконструировать действительную среду обитания человека. Однако Кляйн и Круз-Урибе (1984) считают, что можно сделать жизнеспособные палеологические реконструкции, если сравнивать несколько комплексов окаменелостей с помощью статистических методов, при условии, что качество сохранности костей, так и условия их отложения подобны. Каждая ситуация должна оцениваться с большим тщанием.
Сортировка и идентификация
Остатки животных обычно фрагментарны, являясь частями туш, разделанных на археологическом памятнике или в месте охоты. Какая часть туши переносилась на стоянку, в некоторой степени зависело от размеров животного. Маленького оленя можно было принести на плече целиком. Охотники-собиратели иногда устраивали стоянку на месте убиения большого животного, там они поедали часть туши, а часть засушивали. Однако почти всегда кости, найденные на обитаемых памятниках, были разбиты на осколки. Любое съедобное мясо соскребали с костей, из сухожилий делали ремни, из кожи — одежду, сумки, иногда использовали для жилищ. Съедали даже внутренности. Конечности разбивали, чтобы достать костный мозг. Из некоторых костей делали орудия — наконечники гарпунов и стрел, мотыги (рис. 13.2).
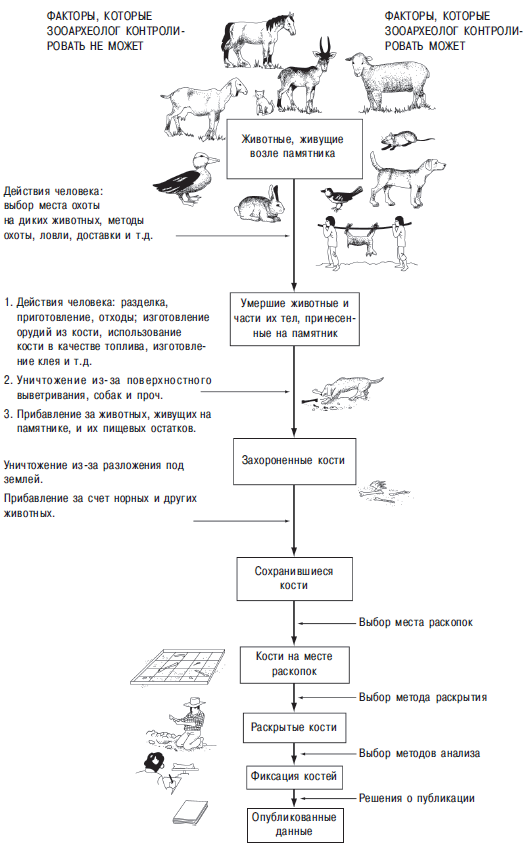
Рис. 13.2. Анализ костей из археологического материала. На этом рисунке показаны некоторые из факторов, которые могут повлиять на данные. Факторы, на которые не могут повлиять археологи, показаны слева, а те, которые археологи могут контролировать, — справа
Было бы ошибкой считать, что по фрагментам костей в археологическом слое можно будет подсчитать точное количество животных, забитых его обитателями, или получить картину окружающей среды во время заселения памятника (Грейсон — Grayson, 1984). Эти кости подвергались разным процессам с момента попадания в археологический слой. Тафономические процессы значительно видоизменяют захороненные кости, кости маленьких животных могут быть вообще разрушены, хотя это нельзя сказать о костях больших. Кроме того, имеются человеческие факторы: люди могли принести дичь издалека или забить всех своих коз прямо на поселении. У нас нет возможности узнать что-либо о ритуальной роли некоторых животных в древних сообществах, какие табу налагались на охоту на одних животных, а на каких — нет. Как уже указывалось, у нас также нет возможности узнать точно сравнительное соотношение различных видов животных в доисторические времена. Конечно, исследователи не могут использовать кости животных из археологических памятников, чтобы отвечать на такие вопросы. Всегда неизвестно различие между тем, что можно было бы назвать собственно «животным» и «археологическим животным», идентифицированным учеными (С. Дэвис — S. J. M. Davis, 1987; Грейсон — Grayson, 1981). Археологическое животное представляет из себя россыпи разбитых человеком костей, которые после этого подвергались разрушительному действию почвы в течение сотен и тысяч лет.
В большинстве случаев идентификация проводится посредством прямого сравнения с известными видами. Это сравнительно легко, и обучиться этому несложно любому человеку с острым глазом (С. Дэвис — S. J. M. Davis, 1987). Но только небольшая пропорция костей в коллекции достаточно полна для этой цели. Рисунок собаки на рис. 13.3 иллюстрирует типичный скелет млекопитающего. Небольшие фрагменты черепа, позвоночник, ребра, лопатки, тазовые кости обычно мало пригодны для того, чтобы отличить домашнее животное от дикого или один вид антилопы от другого. Легко идентифицировать верхнюю и нижнюю челюсти, расположение в них зубов и отдельные зубы, костную сердцевину рога и иногда суставные поверхности длинных костей. Зубы идентифицируют, сравнивая острые выступы на их поверхности с зубами из сравнительных коллекций, тщательно собранных в регионе памятника (рис. 13.4).
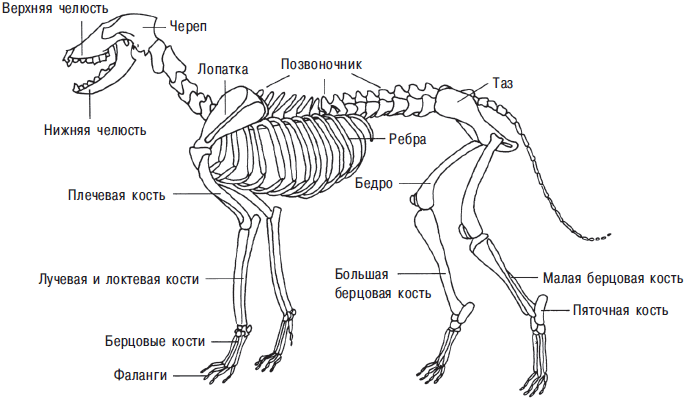
Рис. 13.3. Скелет собаки. Показаны наиболее важные части тела с остеологической точки зрения
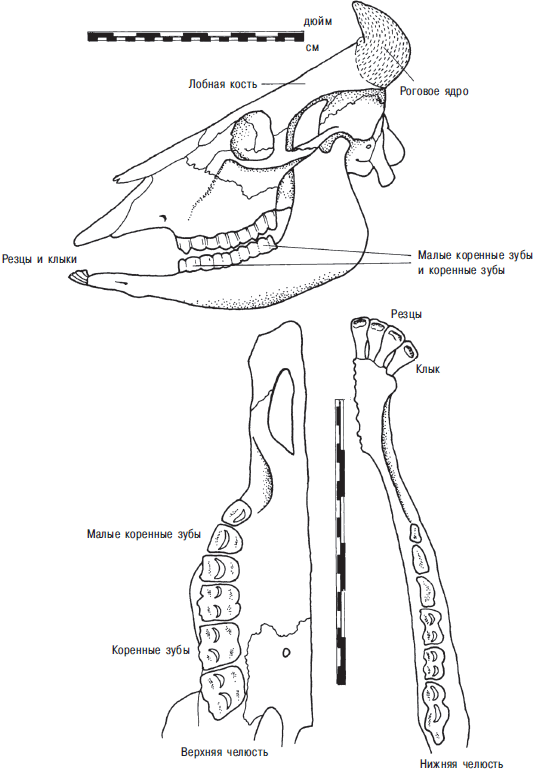
Рис. 13.4. Череп и челюсть домашнего буйвола и наиболее важные остеологические признаки. Одна четвертая реального размера
В некоторых частях мира можно использовать и суставные концы длинных костей, особенно на Юго-Западе Азии или в некоторых частях Северной Америки, где местная фауна млекопитающих довольно мала по числу видов. На юго-западе Азии возможно даже различить домашних и диких животных одного веса по фрагментам длинных костей, при условии, что коллекции достаточно велики и сравнительный материал достаточно полный и в нем представлены все возрасты особей и вариации по размерам самок и самцов. Но в других регионах, например в части Африки южнее Сахары, местная фауна настолько богата и разнообразна и вариации в анатомии скелетов насколько велики, что только сердцевина рога или зубы может помочь различить виды антилоп или дикие и одомашненные формы животного. В заблуждение иногда вводят даже зубы, потому что острые выступы на зубах, например, буйвола и домашнего скота очень похожи и часто единственное различие заключается в меньших размера последнего. Специалисты часто не соглашаются в вопросе о том, в чем состоит определимость кости, поэтому лучше оперировать понятиями разных уровней идентификации, чем просто отвергать возможность определения многих фрагментов. Например, иногда можно идентифицировать фрагмент кости как принадлежащий среднему по размерам плотоядному животному, хотя нельзя утверждать, что он принадлежит волку. Стадия идентификации при анализе кости является самой важной, поскольку при этом нужно ответить на фундаментальные вопросы: это одомашненные или дикие виды? Каково соотношение каждой из групп? Какого рода домашний скот держали обитатели памятника? Были ли у них какие-либо предпочтения в охоте, которые отражались бы в пропорции дичи, обнаруженной в слоях заселения? Все ли дикие виды, характерные для фауны прошлого, существует в данном регионе сегодня?
Сравнение комплексов костей
Зооархеологи Ричард Кляйн и Кэтрин Круз-Урибе (1984) описывают критерии подсчета таксономического обилия для различения реальных комплексов костей и смещенных, то есть полученных в результате пристрастного сбора или под действием других факторов. Они используют те же критерии и для оценки относительного обилия разных видов. Число идентифицированных образцов (ЧИО) — величина количества костей или фрагментов костей от каждого вида в выборке костей. Этот критерий имеет явные недостатки, особенно потому, что он может преувеличивать важность одних видов, костей которых больше, чем других, только потому, что туши тех видов были разделаны более тщательно, чем других. На ЧИО могут повлиять как действия человека, например разделка туш, так и естественные процессы, такие как выветривание. Однако у ЧИО есть определенное важное значение, особенно при использовании его для оценки минимального количества особей, от которых получились идентифицированные кости. Минимальное число особей (МЧО) — величина количества особей, необходимых для того, чтобы получить такое-то число всех идентифицированных костей. Эта величина меньше, чем ЧИО, и часто основана на тщательном подсчете таких индивидуальных частей тела, как пяточные кости. На МЧО не распространяются многие ограничения ЧИО, потому что эта величина является более точной оценкой действительного числа животных. Однако точность зависит от использования специалистами одного и того же метода подсчета МЧО, что часто нарушается (Грейсон — Grayson, 1984).
Вместе взятые ЧИО и МЧО позволяют нам оценивать число животных, наличествующих в костной выборке. Но они являются очень несовершенными методами измерения обилия животных в археологической коллекции, не говоря уже о том, чтобы дать возможность соотнесения костных материалов с популяцией живых животных в прошлом. Кляйн и Круз-Урибе среди других разработали сложные компьютерные программы для того, чтобы преодолеть некоторые из ограничений ЧИО и МЧО, программы, которые выдают базисную информацию, жизненно важную для сравнений выборок между собой.
Видовая структура и культурные изменения
Во время ледникового периода большинство долговременных перемен в видовой структуре животных было вызвано климатическими изменениями, а не культурными. Но некоторые перемены в ней должны и отражать деятельность человека, то, каким образом люди использовали животных (Кляйн и Круз-Урибе — Klein and Cruz-Uribe, 1984). Эти перемены, однако же, очень трудно отличить от тех, что вызваны изменениями в окружающей среде. Одно из таких мест, где стало возможным документально подтвердить такие изменения, является Южная Африка.
Дичь
Хотя перечень дичи и описание повадок животных позволяют понять практику охоты, но во многих случаях содержание этого перечня приобретает особое значение, особенно когда мы хотим понять, почему охотники сосредотачивались на одних видах и явно игнорировали другие.
Табу. Доминирование одного вида дичи могло быть результатом хозяйственной необходимости или удобства или просто делом культурных предпочтений. Многие сообщества ограничивают охоту на некоторых животных или потребление мяса разной дичи по половому признаку. У современного племени Кунг Сэн в районе Добе в Ботсване имеются сложные персональные табу на употребление мяса млекопитающих, зависящие от возраста и пола (Ли — Lee, 1979). Никто не может употреблять в пищу мясо всех 29 видов животных, и у каждого человека свои неповторяющиеся ни у кого табу. Некоторых млекопитающих могут употреблять в пищу все члены племени, но не все части животного. Надзиратели по ритуалам могут наложить и другие ограничения: нельзя есть приматов и некоторых плотоядных. Такие сложные табу повторяются с многочисленными вариациями и в других сообществах охотников-собирателей и земледельцев, и это, несомненно, отражается на пропорции остатков дичи, обнаруживаемых на археологических памятниках.
Примеры специализированной охоты часто встречаются с древних времен, хотя редко можно объяснить причины того или иного предпочтения. Хорошо известно ведение хозяйства на основе охоты на крупную дичь у индейцев прерий (Фрисон — Frison, 1978). Другим фактором, определяющим специализированную охоту, является чрезмерная охота или постепенное вымирание любимых видов. Хорошо известным примером является европейский тур или дикий бык Bos primigenius (рис. 13.5), который являлся главной добычей охотников верхнего палеолита в Западной Европе и на которого охотились в послеледниковые времена и даже после того, как началось производство пищи (Куртен — Kurten, 1968). Последние туры погибли в Польше в 1627 году. По описаниям и изображениям мы знаем, как выглядело это животное. Они были крупными, до двух метров в холке, часто с длинными рогами. Самцы были черного цвета с белой полосой на спине и светлой длинной шерстью между рогов. Немецкие и польские биологи путем длительной селекционной работы успешно воссоздали это животное. На воле воссозданные туры очень темпераменты, свирепы и подвижны. Эти эксперименты дали намного более убедительную реконструкцию одного из самых устрашающих млекопитающих плейстоцена, чем могло бы дать любое количество реконструкций по скелетам или рисункам художников.
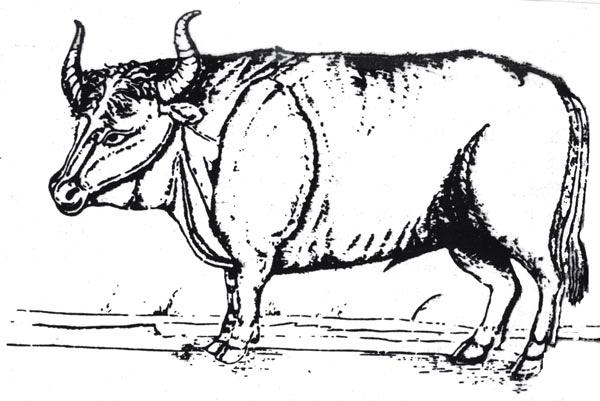
Рис. 13.5. Зубр, как изобразил его С. фон Хербенстайн в 1549 г.
ПРАКТИКА АРХЕОЛОГИИ
ИЗМЕНЕНИЯ В ПРАКТИКЕ ОХОТЫ В ДРЕВНЕЙ ЮЖНОЙ АФРИКЕ
Зооархеолог Ричард Кляйн занимался проблемой соотнесения видовой структуры и культурных изменений, изучая большие выборки фауны из двух прибрежных пещер в Капской провинции, ЮАР. В пещере у реки Клэсис (далее пещера Клэсис) обитали охотники-собиратели среднего каменного века в период между 130 000 и 95 000 лет назад, во время потепления климата, и затем приблизительно до периода 70 000 лет назад, когда климат стал намного холоднее. В более теплые времена море подходило близко к пещере. Многочисленные моллюски, кости тюленей, остатки пингвинов многое рассказали нам о питании людей в этой пещере в средний каменный век. Остатки рыб и морских птиц попадаются редко. Остатки антилопы канна встречаются чаще, чем остатки другие млекопитающих, например более чем в 2 раза чаще, чем остатки буйвола. Остатки других наземных млекопитающих принадлежат видам, распространенным в современные исторические времена. В противоположность этому, в близлежащей пещере у залива Нельсона (далее пещера Нельсона) имеются следы обитания человека в позднем каменном веке, приблизительно 20 000 лет назад. В тот период последнего оледенения море находилось уже в нескольких километрах от пещеры. В этой пещере было много остатков летающих морских птиц и рыб, а остатков антилопы канны — только третья часть, столько же, сколько и буйвола.
Кляйн указывает также на то, что наборы орудий были совершенно разными в этих пещерах. Люди среднего каменного века из пещеры у реки Клэсис использовали большие орудия-отщепы и копья, а у охотников из пещеры Нельсона были луки и стрелы и большой набор маленьких каменных орудий и артефактов из кости, некоторые из них были сделаны для особых целей, например для ловли птиц и рыболовства. Эти инновации позволяли охотникам позднего каменного века убивать более опасных и осторожных животных с большей частотой. Таким образом, причина того, что людям среднего каменного века чаще попадались антилопы канна, лежит не в том, что она была более распространенной, а в том, что более сложную дичь убивали реже. Все указывает на то, что племена Клэсис поведенчески (behaviorally) были менее продвинутыми, чем люди из пещеры Нельсона (Klein and Cruz-Uribe, 1984).
Кляйн соединяет некоторые другие сведения о фауне уже с климатическими данными. Памятник у реки Клэсис содержит остатки черепах и моллюсков блюдечек гораздо больших размеров, чем в последующие времена, как если бы этим существам позволяли гораздо дольше расти. Эти факты говорят о меньшем давлении на популяции черепах и моллюсков со стороны немногочисленного населения людей до того, как появились более технологически развитые племена.
Изменения в охоте. В последнее время охота сильно изменилась. Ричард Ли (Richard Lee, 1979) записал рассказы стариков племени Сэн об охоте в прежние времена. Тогда в центральной части Ботсваны было больше дичи и больше охотников. Их предки большими группами охотились на буйволов, жирафов и слонов. Сегодня у племени преобладающим видом хозяйства является собирание, дополнением является употребление мяса 29 видов млекопитающих, в основном таких, у которых с одной туши можно получить сравнительно больше мяса. Охота ведется методом преследования, главным источником мяса является африканская свинья — бородавочник и мелкая дичь. Такие изменения в охоте являются прямым результатом импорта ружей и первых охотничьих сафари, которые уничтожили замечательную фауну Африки в течение жизни трех поколений.
Сезонные занятия. Многие доисторические охотники-собиратели и земледельцы, как и их современные собратья, вели жизнь, основанную на смене времен года, их деятельность по жизнеобеспечению менялась посезонно. На северо-западном побережье Тихого океана, когда летом лосось начинал двигаться вверх по течению рек, возле них собирались индейцы, ловили тысячи рыб и засушивали их на зиму. В начале сухого времени года в Центральной Африке было изобилие диких плодов, которые составляли важную часть рациона древних земледельцев 1500 лет назад. Как же археологи изучают сезонные виды деятельности и реконструируют «экономические времена года»?
Любые аспекты жизни древних охотников-собирателей были связаны со сменой времен года. В долгие зимние месяцы племена северо-западных индейцев занимались сложными ритуалами. Жизнь племени животноводов хо-хо в районе мыса Доброй Надежды резко изменялась во время сухих или дождливых времен года (Элфик — Elphick, 1977). В течение сухих месяцев они собирались у нескольких постоянных источников воды и возле невысыхающих рек. При наступлении дождей они выгоняли свой скот на близлежащие земли, насыщая свои стада влагой из стоячих вод, оставшихся после ливней. Как же археологи изучают сезонность? Успех приносили множество методов (Монкс — Monks, 1981). В самых простых из них с помощью костей и остатков растений определяют, когда на памятнике были люди. Например, на памятнике возрастом 1000 лет в заливе Сан-Франциско люди бывали каждый год приблизительно 28 июня, когда бакланы были еще молодыми (Хоуард — Howard, 1929) (обсуждение вопроса о птицах см. далее в этой главе). Наличие костей трески в древних норвежских памятниках говорит о том, что они были обитаемы в течение зимы и ранней весны, оптимального времени для сушки рыбы. Этот вид анализа хорош, при условии, что повадки животных или доступность растений, рассматриваемых в данной ситуации, хорошо известны и не изменились со временем. Многие растения доступны в течение большей части года, но пригодны к употреблению в пищу только в течение нескольких недель.
Знание экологии как животных, так и растений необходимо, так как «график» использования ресурсов, хотя, быть может, и не точный, но, безусловно, являлся важнейшим фактором в жизни древних сообществ (см. вставку «Практика археологии»). Некоторые животные, такие как олень, сравнительно безразличны к сезонным переменам, но люди использовали их по-разному в разное время года. Например, индейцы племени сэлиш с северо-западного побережья Тихого океана весной брали самцов, а осенью — самок (Монкс — Monks, 1981).
Кроме того, имеются физиологические явления в жизни животного, по которым археологи могут определить сезон встречи с ним. В течение XV века н. э. группа охотников Великих равнин регулярно охотились на бизонов около источника воды у Гэрнсея, штат Нью-Мексико (Спет — Speth, 1983). Джон Спет (John Speth) проанализировал части тел на месте забоя и обнаружил, что охотники весной, в сезон охоты, явно предпочитали самцов. Те, кто разделывал туши, оставили на памятнике такие части тела, которые давали мало мяса, — головы и верхние участки шеи, а тех частей, которые давали много мяса, жира и костного мозга, было мало. Также из самцов больше, чем из самок, брали костей для последующего использования. Спет считает, что охотники предпочитали самцов, потому что после зимы они были в лучшем состоянии и их мясо было жирнее.
Иногда на сезонные занятия может указывать возраст животных. По мере взросления животного эпифизы на конце костей конечностей медленно соединяются с основным телом кости, и эти места полностью окостеневают. При их изучении можно определить общий возраст животных, скажем, на стоянке охотников, но такие факторы, как питание, даже кастрация одомашненных животных, могут повлиять на скорость этого процесса. Некоторые виды, утки например, взрослеют намного быстрее, чем олени. Понятно, что при таком подходе необходимы знания о возрастных изменениях в суставах.
Всем известно, что при взрослении выпадают молочные зубы, у людей часто бывают проблемы с зубами мудрости. Зубы относятся к настолько прочным остаткам животных, что многие археологи пытались использовать их для определения возраста диких и домашних животных. Достаточно легко изучать выпадение зуба из целых и даже фрагментарных челюстей, и это было сделано на примере домашних овец, коз и диких оленей. И опять факторы питания, одомашнивания могут повлиять на скорость выпадения зубов, и скорость износа зубов может сильно различаться у разных популяций (Монкс — Monks, 1981).
ПРАКТИКА АРХЕОЛОГИИ
ОКРУЖАЮЩАЯ СРЕДА И СЕЗОННОСТЬ НА ПАМЯТНИКЕ СТАР КАРР, АНГЛИЯ
Памятник Стар Карр на северо-западе Англии был заселен небольшой группой охотников-собирателей каменного века приблизительно в 8500 году до н. э. Это крохотное поселение, в котором полвека назад нашли хорошо сохранившиеся редкие артефакты из кости и дерева, известно во всем мире тем, что оно дало замечательно полную картину жизни в Северной Европе сразу же после последнего ледникового периода. Между 1949 и 1951 годами археолог Грэхэм Кларк (Grahame Clark, 1954) из Кембриджского университета обнаружил там маленькую платформу из березового дерева, усыпанную фрагментами орудий из камня, артефактов из кости и дерева и множеством остатков пищи. Используя тщательно зафиксированные подсчеты артефактов, кости животных, анализ пыльцы и разного рода сложные способы идентификации, а также щедрую долю традиционного европейского фольклора, Кларк реконструировал небольшую охотничью стоянку в зарослях камыша у озера. Анализ пыльцы показал, что Стар Карр существовал в то время, когда березовые леса впервые распространились по северу Британии, а большая часть южной части Северного моря была еще сушей. Кларк со своими коллегами утверждали, что памятник был обитаем зимой, свидетельством этому служили рога марала. Кларк проанализировал методы изготовления наконечников копий из кости, связал технологию изготовления каменных орудий с теми, что были сделаны в Скандинавии в то же время, и описал замечательные серии орудий из кости и дерева, включая мотыги из рога лося (одна из них была с остатком деревянной ручки), цельное деревянное весло для каноэ, шила и даже кусочки коры и мха для розжига огня (рис. 13.6).
В течение полувека памятник Стар Карр стал важным полигоном для апробации новых идей в отношении сообществ охотников-собирателей. Археологи Пол Мелларз и Петра Дарк (Paul Mellars and Petra Dark, 1999) недавно завершили 12-летние чрезвычайно избирательные палеоэкологические и археологические исследования на памятнике с использованием всех ресурсов современной науки для повторной интерпретации памятника. Когда Кларк первоначально раскапывал Стар Карр, он сосредоточился на небольшом заболоченном участке в овраге. После трех сезонов он интерпретировал памятник как небольшое поселение, возможно нерегулярно используемое четырьмя или пятью семьями. Новые расширенные раскопки распространились на более сухие участки и показали, что памятник намного больше, чем предполагал Кларк. С помощью полевых съемок и осторожного рытья пробных шурфов археологи обнаружили россыпи кремниевых артефактов на расстоянии 12 метров от берега древнего озера. Тщательно изучая первоначальную топографию памятника, Мелларз и Дарк со своими коллегами обнаружили заполненный глиной канал, который когда-то проходил через центр памятника, отделяя заболоченный участок, который изучал Кларк, от более сухих участков.
Кларк утверждал, что обитатели Стар Карра мало влияли на среду обитания. Дарк смогла использовать микроскопы с большей разрешающей способностью для изучения распределения угольных частиц, связанных с новом массивом радиоуглеродных дат, полученных с помощью ускоряющей масс-спектрометрии. Она показала, что имел место инициальный период интенсивного отложения угля, который длился около 80 лет. Затем следовало 100 лет слабой активности, за которыми последовало достаточно длительное отложение в течение еще 130 лет. Ботаник Йон Хэтер идентифицировал угли как прибрежный камыш, сожженный в сухом виде между осенью и весной, когда начинается новый рост. Мелларз и Дарк считают, что люди неоднократно сжигали камыш, главным образом потому, что образцы угля показывают, что пожары были локализованы у памятника, как будто огонь находился под контролем. Такие пожары могли дать лучший обзор озера и прилегающей территории, а также удобное место для причаливания каноэ, а новая растительность привлекала бы кормящихся животных.
Первоначальный отчет Кларка описывал Стар Карр как зимнее поселение. Теперь же рентгеновский анализ невыпавших из челюстей зубов оленя и сравнение с современными образцами позволили идентифицировать множество 10- и 11-месячных животных, которые были бы забиты в марте или апреле (Р. Картер — R. Carter, 1998). Эти новые данные о сезонности согласовываются с обнаружением плотно скрученных стеблей камыша, сожженного в начале роста между мартом и апрелем, и чешуйками почек осины, которые датируются тем же временем года. Стар Карр не является зимним поселением, в нем жили с марта по июнь или начало июля.
Рис. 13.6. Мотыга из рога лося из Стар Карра, Англия. (Две трети от реального размера)
Интерпретация сезонных занятий в значительной мере зависит от этнографических аналогий. Классическим примером является дикая пшеница. Ботаник Гордон Хиллман изучал сбор дикой пшеницы на юго-западе Азии и показал, что сборщики должны очень точно рассчитывать время сбора урожая. Это нужно было сделать до того, как колосья осыплются или зерна употребят в пищу птицы или животные (Хиллман и Дэвис — Hillman and Davis, 1990). Разумно предположить, что подобное точное планирование было необходимо и в доисторические времена. Эта аналогия позволила археологам Юго-Западной Азии интерпретировать сезонные занятия на памятниках в Сирии и других местах.
Изучая не только крупных млекопитающих и большие растительные остатки, но также и мельчайших моллюсков и чешую рыб, можно уточнить рамки сезонных занятий до удивительно узких границ.
Домашние животные
Почти все домашние животные произошли от диких видов, склонных к общению с человеком (Клаттон-Брок — Clutton-Brock, 1981, 1989). Нельзя сказать, что все домашние животные происходят из одной части света, их одомашнивали в их естественной среде обитания. Ученые предполагают, что одомашнивание диких животных совершается при достижении определенного культурного уровня. Кажется, что везде одомашнивание начиналось тогда, когда растущее население нуждалось в более регулярном снабжении питанием, когда нужно было кормить большие группы людей. Одомашнивание зависит от этого условия и является предварительным требованием для дальнейшего роста населения.
У диких животных отсутствуют многие качества, которые ценят в их домашних аналогах. Так, у диких овец обильная шерсть, но она качественно не такая, как у домашних, которая пригодна для прядения. Дикие козы и буйволицы дают достаточно молока для их детенышей, но не в том количестве, что необходимо для человека. Во время одомашнивания люди развивали в животных нужные им свойства, произошедшие изменения часто делали животных непригодными для выживания в дикой природе.
История домашних видов основана на фрагментах костей животных, найденных в слоях многочисленных пещер, укрытий и на открытых памятниках (Клаттон-Брок — Clutton-Brock, 1989). Остеологическое изучение диких и домашних животных сдерживает как фрагментарность костей на большинстве памятников, так и более широкие границы возрастной изменчивости домашних животных по сравнению с дикими (Цедер и Гессе — Zeder and Hesse, 2000; Цедер — Zeder et al., 2002). Тем не менее на ряде памятников были получены данные о постепенных остеологических изменениях в сторону домашних животных. Если сравнивать кости дикого вида каких-нибудь доисторических одомашненных животных с костями этого домашнего животного во времени, то диапазон изменений размеров сначала возрастает, затем, в конце концов, выбор делается в пользу меньших животных, и изменения в размерах также становятся меньше. Этот переход плавный, и потому крайне трудно идентифицировать домашнее или дикое животное по отдельным костям или маленьким коллекциям.
Кости домашних животных показывают, что диким видам была присуща высокая степень адаптивности. Люди сочли нужным изменять размеры и свойства животных в соответствии со своими нуждами, что отразилось на скелетных останках животных. Со времен начала одомашнивания животных были выведены разные породы скота, овец и других домашних животных.
Забой и разделка туш
Некоторое представление об эксплуатации диких и домашних животных можно получить, изучая не только сами кости животных, но и их частотность и распределение в почве.
Пол, возраст и забой. Ясно, что определение пола животного и возраста, в котором его забили, помогает при изучении охоты или методов содержания стад теми людьми, которые занимались забоем. У археологов есть много методов для установления пола и возраста животных по фрагментам костей (С. Дэвис — S. J. M. Davis, 1987).
Мужские и женские особи многих млекопитающих значительно отличаются по размерам и строению. У жеребцов, например, имеются клыки, а у кобыл — нет. У людей строение таза женщин отличается от мужского, что связано с деторождением. Мы можем оценить соотношение особей мужского и женского рода на таких памятниках, как место забоя бизонов Гарнсей, сравнивая количество частей туш самцов и самок, так как для этого вида разница между ними известна. Подобный анализ намного труднее проводить, если мало известно о различиях в размерах или когда кости очень фрагментарные. Зооархеологи используют множество измерений костей для различения полов, но такой подход чреват статистическими и практическими трудностями; он хорошо срабатывает только при целых костях. Но даже и тогда возможно идентифицировать лишь распределение различных измерений (размеров), которые могут отражать или не отражать различия между полами.
В каком возрасте забили этот скот? Отдавали предпочтение обитатели поселения мясу незрелых диких овец или взрослых? Для многих памятников это важные вопросы. Для того чтобы ответить на них, исследователи должны установить возраст животных в выборке на момент их смерти. Обычно для этого используют зубы и эпифизы концов конечностей. Почти у всех млекопитающих кости, на которых эпифизы не приросли, принадлежат молодым особям. Этот факт позволяет нам говорить о двух классах: незрелых и взрослых животных. Если мы знаем возраст, в котором срастаются эпифизы, как иногда в случае с такими видами, как крупный рогатый скот, можно вводить дополнительные классы. К сожалению, эпифизарное сращение является слишком общим методом для получения тех данных, в которых нуждаются археологи.
К счастью, зубы верхней или нижней челюстей позволяют более точно определить возраст животных. Зубы являются непрерывной нитью, по которой можно определить жизнь от рождения до старости. Цельные верхние и нижние челюсти позволяют нам изучить незрелые и зрелые зубы по мере их выпадения, таким образом мы можем определить не только молодых животных, но и старых.
Отдельные зубы также могут быть источником информации о возрасте животного. Некоторые биологи используют кольца роста на зубах, но этот метод до сих пор носит экспериментальный характер. Многообещающим методом является измерение высоты коронки зуба. Ричард Кляйн, специалист по костям африканских животных, измерил высоту коронок зубов млекопитающих каменного века, найденных в пещерах у реки Клэсис и в заливе Нельсона в Капской провинции ЮАР. Разделенные на две группы, замеры зубов дают интересные общие представления об охоте в средний и поздний периоды каменного века в этом регионе (Кляйн — Klein, 1977). Кляйн сравнил распределения смертности капских буйволов и других крупных и средних видов с кривыми смертности их современных популяций. Он идентифицировал два основных распределения для костей каменного века (Кляйн и Круз-Урибе — Klein and Cruz-Uribe, 1983). В катастрофическом одномоментном возрастном профиле меньше старых особей. Это нормальное распределение живущих ныне популяций копытных (рис. 13.7, столбец слева) и обычно обнаруживается в местах массового забоя, при загоне стада в болото или в овраг с крутого обрыва, а также при гибели всей популяций по естественным причинам. Разреженный возрастной профиль (рис. 13.7, столбец справа) показывает недостаточную долю животных в своем лучшем возрасте относительно их количество в живущих популяциях, но молодые и старые особи представлены избыточно. Считается, что такой профиль является результатом питания падалью или простой охоты копьем.
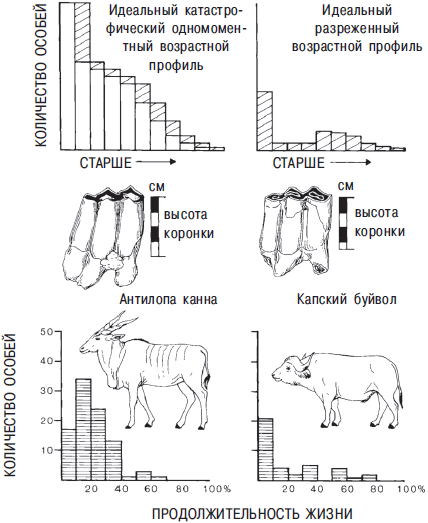
Рис. 13.7. Схематичные данные смертности современных популяций животный на основе молярных коронок двух распространенных африканских млекопитающих — антилопы канна и капского буйвола: левый столбец — схематический катастрофический возрастной профиль; правый столбец — схематический разреженный возрастной профиль
Кляйн обнаружил, что возрастные распределения капских буйволов на обоих памятниках близки к распределениям, наблюдаемым для современных буйволов, которых убивают львы, и это может быть потому, что молодые и старые самцы являются уязвимой мишенью из-за своей изолированности от больших стад зрелых грозных животных. Поэтому он утверждал, что охотники обеих пещер каменного века стабильно и в течение длительного времени использовали популяции буйволов. Распределение антилопы канна и гибридов бубала (меньшие стадные антилопы) намного больше похоже на катастрофический профиль. Кляйн предполагает, что они подобны потому, что на эти виды охотились массовым загоном, как на бизонов на американском плато Великих равнин. Таким образом, целые популяции могли быть убиты за один раз. Возрастные распределения могут отражать любые другие виды активности. На памятнике Стар Карр на северо-востоке Англии нет молодых маралов. Большинству животных было три или четыре года, неопытные подростки погибали, когда оставляли матерей (Легге и Роули-Конуи — Legge and Rowley-Conwy, 1988).
На охоту и забой животных воздействуют разного рода мелкие факторы, многие из которых описаны Льюисом Бинфордом (Lewis Binford (1978, 1981b). Изучая практику охоты племен нунамиутов на Аляске, он обнаружил, что забой охотниками животных является частью намного более обширной стратегии жизнеобеспечения. Нунамиуты в течение большей части года в значительной степени полагаются на заготовленное мясо, поэтому при охоте ориентируются и на цели заготовления, и на многие другие. Осенью они могут охотиться на оленят для получения кожи для зимней одежды, причем головы и языки этих животных дают пищу тем, кто обрабатывает кожи. Бинфорд подчеркивает, что трудно интерпретировать структуры забоя без точного понимания культурной системы, частью которой является охота.
Домашние животные являются контролируемым источником мяса, и здесь применяются совершенно иные критерии отбора. В более развитых земледельческих сообществах крупный рогатый скот или лошадей могли держать до старости в качестве тягловых животных, лишних мужских особей кастрировали, а женских поддерживали до тех пор, пока они не переставали давать молоко, приплод или не приносили больше пользы при вспашке земли. Если даже не держали ездовых или рабочих животных, проблема лишних мужских особей сохранялась. Этот излишек являлся обильным источником мяса, и этих животных часто забивали в раннем взрослом возрасте. Во многих традиционных обществах скот являлся мерилом благосостояния, как, впрочем, и сегодня, и скот в них забивают по особым случаям — на свадьбах или похоронах. Таким образом, потреблялись излишки стада и удовлетворялись запросы владельца стада.
Забой скота. Фрагменты костей на уровне заселения являются конечным продуктом забоя, разделки и потребления домашних или диких животных. Для того, чтобы понять этот процесс, нужно изучить сочленение костей животных на тех уровнях, где они были найдены, или должен быть тщательно изучен анатомический состав костей. На памятнике Олсен-Чуббок в Колорадо факты свидетельствуют о забое стада бизонов. Охотники устроили лагерь неподалеку, где снимали кожу и мясо с туш и, возможно, засушивали излишки мяса для потребления в дальнейшем. Разделочные инструменты были найдены в прямом комплексе с костями, таким образом, на этих раскопках навсегда сохранился «момент» разделки туш (Уит — Wheat, 1972).
Интерпретация методов разделки является сложным делом, потому что многие факторы повлияли на то, как расчленяли туши. Племя индейцев нунамиутов в основном полагалось на запасенное мясо, и то, как они расчленяли оленей, зависело от того, сколько нужно было сохранить мяса, от выхода мяса от разных частей тела, от отдаленности основной стоянки. На любом памятнике-стоянке количество найденных костей зависит от размеров животных: тела коз, куриц или мелкой дичи можно было принести целиком, но туши больших животных доставляли частями. Иногда животных с большим выходом мяса употребляли в пищу там, где их убивали, не оставляя ни кусочка мяса или внутренностей. Интерпретация очень сложна даже в отношении индексов МЧО и ЧИО.
И вновь задача заключается в установлении значения археологических распределений для понимания деятельности человека. Насколько это сложно в контексте разделки мяса, можно судить по комментариям Бинфорда (Binford, 1978) о том, что критериями для выбора мяса индейцами нунамиутами являются количество годного к употреблению мяса, время, необходимое для его обработки, и качество мяса.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
ИГРА В КОСТИ
ИГРА В КОСТИ Поднимаясь по лестнице в комнату, которую он занимал на чердаке, Танге еще. издали услышал грубый голос, выкрикивающий двусмысленные слова немецкой солдатской песни. Чтобы поскорей прервать это непристойное пение, капеллан торопливо вошел в распахнутую
Черепа и кости
Черепа и кости Из черепов вождей враждебных кланов скифы изготовляли ритуальные чаши, из которых пили ритуальное вино в особых случаях, когда надо было привлечь душу побежденного противника себе в союзники. Впоследствии гунны переняли у саков этот обычай и стали
Изрезанные кости из Италии
Изрезанные кости из Италии Образцы, изрезанные способом, сходным с образцами из Сант-Преста, были найдены Ж. Деснойерсом в коллекции костей, собранных в долине реки Арно в Италии. Кости с желобками принадлежали тем же типам животных, что найдены и в Сант-Престе, в том числе
Резчики по слоновой кости
Резчики по слоновой кости В мастерской микенского резчика на самом светлом месте лежали семь бронзовых инструментов: маленькая пилка, три резца (один — с изогнутым лезвием), напильник, щербатка (своего рода цилиндрическая терка), бурав. Режущая часть делалась из осколка
«Череп и кости»
«Череп и кости» «Чаю воскресения мертвых и жизни будущего века. Аминь». Надпись на «знамени смерти» генерала Бакланова Головные уборы частей СС особого назначения также весьма напоминали армейские. Но, в отличие от головных уборов солдат германского вермахта, головные
Поддельные кости
Поддельные кости Одним из доказательств древности происхождения человека, по мнению Кремо, служат обработанные или сломанные кости животных. Первые подобные находки были сделаны в XIX веке, и они продолжаются до сих пор. Сторонники теории Дарвина утверждают, что эти кости
5. Яркие желтые кости
5. Яркие желтые кости ПЕРЕСЕКАЯ ВЫСОКИЕ РАВНИНЫРано выехав из мотеля, к 6 часам утра я был на дороге к Блэкуотер-Дро, знаменитому месту раскопок эпохи кловис. Он располагался около Кловис, Нью-Мексико, где был найден первый наконечник кловис. Солнце поднималось медленно,
Плечи из слоновой кости
Плечи из слоновой кости «В самый день его усыновления юному Веру (Марку Аврелию), — пишет его биограф, известный под именем Юлия Капитолина, — почудилось, будто плечи его из слоновой кости, будто он попробовал, какой груз они могут вынести, и убедился, что плечи его стали
5. Яркие желтые кости
5. Яркие желтые кости ПЕРЕСЕКАЯ ВЫСОКИЕ РАВНИНЫРано выехав из мотеля, к 6 часам утра я был на дороге к Блэкуотер-Дро, знаменитому месту раскопок эпохи кловис. Он располагался около Кловис, Нью-Мексико, где был найден первый наконечник кловис. Солнце поднималось медленно,
Часть III Варяжские «кости»
Часть III Варяжские «кости» Введение – Господа, господа! Прошу вас, не устраивайте ажиотацию. Все билеты проданы и войти вам совершенно нет возможности.Смотритель Броницкий еле отбивался от наседающей толпы, призывно поглядывая на стоявшего поодаль пристава Новикова.
Богатырские кости
Богатырские кости Древние, допотопные кости мамонтов большей частью почитались у нас костьми богатырскими. И там, и сям рассказывали о богатырях гигантах. В округе города Переславль-Залесского один помещик употреблял плоскую мамонтову кость вместо печной заслонки,
Мордовские кости
Мордовские кости Верстах в восьми от Нижнего Новгорода, в сторону Арзамасского тракта, между деревней Щербинки (она же Новая), стоящей на самом тракте, и деревней Ляхово в настоящее время пролегает сухой дол — когда-то пойма, теперь пересохшая в этом месте, у бывшего
Люди «белой кости» и люди «черной кости»
Люди «белой кости» и люди «черной кости» Традиционное казахское общество имело строго иерархическое устройство. Идея наследственной аристократии выражалась резко, так что аристократия и представители так называемых «святых родов» были четко отделены от
Кости из песчаного карьера
Кости из песчаного карьера Было утро выходного дня.Не спеша, дядя Ваня впрягает Монголку в повозку. Сережка-ремесленник в кустах на тропинке поджидает Валечку-гимнастку. Детвора, макая соломинки в мыльную воду, что в банке консервной, пускает пузыри.В это время по улице
Бог снова бросает кости
Бог снова бросает кости – Что же касается путей в будущее, то здесь я согласен с Гераклитом, провозгласившим: «Борьба – отец всего», – продолжает профессор. – Не думаю, что послекапиталистический мир будет таким единым. Посмотрите, как обстояло дело в таком значительно
Кости динозавра
Кости динозавра Садгери – это небольшое село выше Боржоми. Там древний храм – базилика X или XI века. Летом в этом селе останавливаются отдыхающие, в основном с детьми. В тот день я только вернулся из Тбилиси, и матушка Анна радостно сказала при встрече:– Благословите,