АНТРОПОГЕНЕЗ
АНТРОПОГЕНЕЗ
Примерно два с половиной миллиона лет назад на планете Земля появился новый биологический вид, отличавшийся от всех своих предшественников тем, что обладал свойствами выживать, опираясь в большей степени на интеллект, чем просто на физические способности организма. Человек вышел из природного царства «без фанфар и барабанного боя», но событие это стало настолько значимым для самой планеты, что, осмысливая глобальную историю Земли, ученые начали вести с этого момента отсчет продолжающейся и поныне новой геологической эпохи, названной антропогеном.
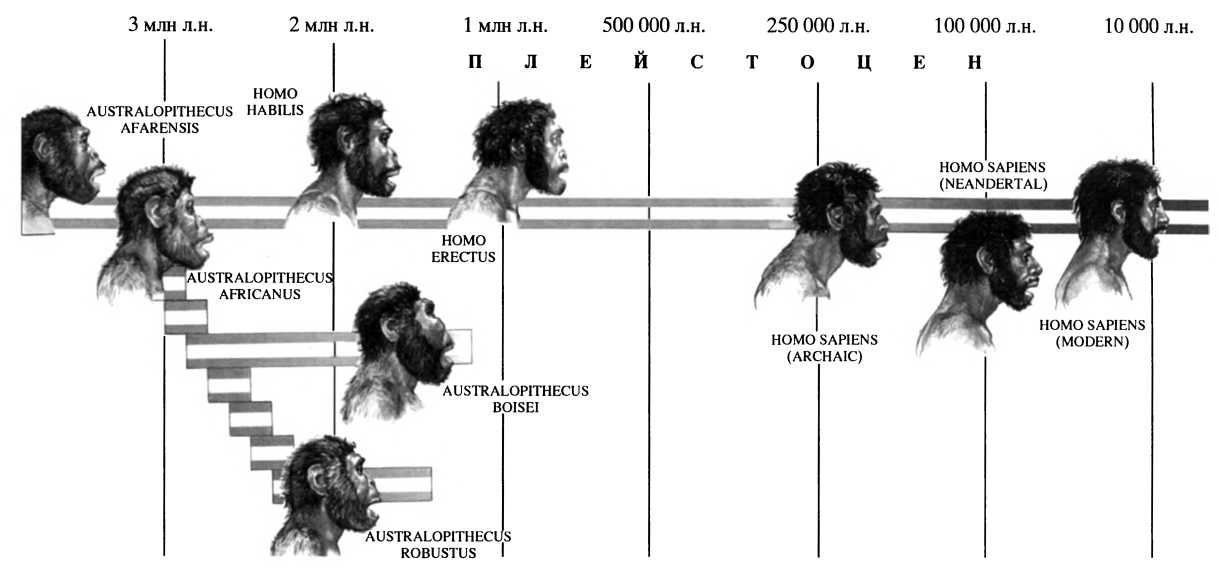
Схема магистральной линии антропогенеза согласно наиболее распространенной научной версии
Проблема происхождения человека в подлинно научном виде впервые была сформулирована в рамках эволюционного учения Ч. Дарвина. В двух его работах: «Происхождение видов путем естественного отбора» (1859) и «Происхождение человека и половой отбор» (1871) возникновение человеческого рода было представлено в качестве органической части общей биологической эволюции. Принципиальные заключения сводились к тому, что человек имеет единого с современными антропоморфными обезьянами предка, становление человека происходило, скорее всего, на африканском континенте и в основе трансформации животного предка в человека решающую роль играли механизмы, ориентированные на индивидуальный и популяционный отбор, обеспечивающий наибольший успех в выживании вида.
Современные микробиологические исследования подтвердили выводы об африканской прародине человека и о филогенетическом родстве современного человека с высшими приматами. Из четырех современных видов антропоморфных обезьян — гориллы, гиббона, орангутана и шимпанзе — по генетическим характеристикам к человеку ближе всего оказалась одна из разновидностей шимпанзе. Поскольку шимпанзе, как и горилла, обитает только на африканском континенте, то ясно, что его общий с человеком далекий предок тоже должен был иметь местные корни.
Что же послужило причиной, пусковым механизмом для трансформации в общем-то благополучного сообщества человекообразных обезьян в семейство гоминид? На логичное объяснение этого процесса претендует гипотеза экологической катастрофы. Согласно ей примерно 5–6 млн лет назад резкие геологические, геоморфологические и климатические изменения привели к существенной перестройке ландшафтов в Африке. В восточной части континента произошло сильное сокращение или даже исчезновение лесного покрова, сопровождавшееся возникновением естественных барьеров по линии геологических разломов, отмеченных в настоящее время цепочкой Великих восточноафриканских озер. Обитавшие здесь человекообразные обезьяны постепенно лишались привычной среды обитания. Часть их могла мигрировать, другая, возможно, вымерла, а какая-то под давлением указанных факторов оказалась способной к усвоению новых поведенческих стереотипов. В западной же части Африки, которой не коснулась происшедшая деградация лесов, они продолжали и продолжают свое существование до наших дней.
Согласно приведенным обобщениям, с указанного выше хронологического рубежа активизируются следующие факторы эволюции: чисто биологические (половой отбор), приспособительные (прямохождение, утеря специализации, смена пищевого состава, орудийная деятельность) и поведенческие (изменение системы иерархии и структурирования сообщества). В дальнейшем уже только количественного развития этих взаимообусловленных факторов оказалось достаточно для того, чтобы сформировался новый социально-биологический феномен природы — человек. Археологически его появление фиксируется, по современным данным, по крайней мере около 2,5 млн лет назад. Бесспорно, с этого времени (а скорее всего, еще раньше) начинается история человека и его культуры.
Спустя почти полтора века после появления дарвиновской теории в постановке конкретных вопросов, связанных с возникновением человека, естественно, произошли изменения. В настоящее время теория антропогенеза предстает как результат синтеза комплексных данных, предоставляемых археологией, палеоантропологией, культурологией, этологией, молекулярной биологией. Новейшие исследования в данной области опираются на солидную базу полноценных источников и обеспечиваются возможностями современных методико-методологических подходов различных научных дисциплин. Изучение рассматриваемой проблемы в наши дни основывается на учете трех взаимосвязанных аспектов: археологического, антропологического и философского. Археологический аспект призван раскрыть вопросы времени и места возникновения человека. Антропологический служит реконструкции морфологического развития при переходе от прегоминид к гоминидам и объяснению биологической основы этого процесса. Философский аспект проблемы состоит в формулировании наиболее адекватного определения человека как существа не только биологического, но и социального.
Критерии. Кажущийся очевидным на современном бытовом уровне вопрос об отличии человека от животного не так прост, когда исследователь оказывается перед проблемой определения эволюционного статуса древнейших представителей человеческого рода. Попытки установить один, два или даже комплекс чисто антропологических универсальных критериев человека («гоминидная триада»), например, таких как определенный объем мозга, прямохождение, развитость кисти руки, не оказались результативными. Зачастую с основывающимися на этих показателях определениями входили в противоречие археологические данные, характеризующие уровень материальной культуры, которая сопутствовала тем или иным палеоантропологическим находкам.
Археологические и антропологические данные составляют основу для установления философского критерия человека применительно к начальным этапам антропогенеза. Определяется этот критерий как способность к труду. Инстинктивные, животнообразные формы орудийной деятельности свойственны и для животных. Но труд человека, как сознательная, систематическая, целенаправленная и разнообразная деятельность, направленная на выживание и воспроизводство, имеет свою специфику. Последняя заключается в существовании такого процесса производства, который включает в качестве обязательного компонента средства производства, какими бы примитивными они ни были. Для каменного века таковыми являются, например, изделия, представляющие собой промежуточные средства для изготовления законченных орудий.
Исходя из приведенного определения и основываясь на археологических находках, возникновение человека относят ко времени не позднее 1,8 млн лет назад. Именно так датируется Homo habilis («человек умелый») — первый представитель рода Homo, костные остатки которого были обнаружены при раскопках древнепалеолитических стоянок Восточной Африки (стоянки ущелья Олдувай в Танзании). Но в действительности человек, по-видимому, возник существенно раньше. Об этом свидетельствует возраст первых орудий труда (не менее 2,4 млн лет назад), происходящих из памятников на территории Кении (стоянка Кооби-Фора) и Эфиопии (стоянка Гона). Неопровержимых данных для заключений о том, кто был изготовителем этих орудий, пока не имеется. С известной долей вероятности их изготовителем можно считать более раннюю форму хабилиса, например Homo rudolfensis. Некоторые исследователи не исключают и того, что их изготовляли австралопитеки, но доказательств этому не находится. Поскольку австралопитеки возникают задолго до появления первых достоверных каменных орудий и морфологически сильно отличаются от Homo habilis, то их не относят к человеку, а выделяют в отдельный род, объединяемый вместе с родом Homo в семейство гоминид. Говоря о вероятной древности человека, некоторые антропологи обосновывают гипотезу о расхождении человека и австралопитеков на пути их эволюции от общего предка ранее 5 млн лет назад.
Таким образом, в традиционной биологической систематике, основанной на морфологических (но не генетических) критериях, современный человек предстает как вид Homo sapiens рода Homo семейства Hominidae. Путь от прегоминид к человеку занял несколько миллионов лет.
Наиболее близкими предшественниками человека в морфологическом и хронологическом отношениях являлись представители рода австралопитековых (Australopithecus). Интерес к австралопитековым всегда был высоким, главным образом, благодаря тому, что их изучение имеет отношение к проблеме антропогенеза. Некоторые из видов австралопитеков впоследствии являлись и современниками раннего человека. Род Australopithecus включает около десятка видов. Территория их возникновения и распространения — Восточная, Южная и Центральная Африка.
Исследователи подразделяют австралопитековых на ранних (7? — 4 млн лет назад), грацильных (4? — 2,5 млн лет назад) и массивных (2,5 млн лет назад). С точки зрения проблемы антропогенеза наиболее интересной считается группа грацильных. Признаков, свойственных человеку, в скелетных остатках этой разновидности выявляют больше, чем у других их собратьев. С другой стороны, находки костей грацильных австралопитеков обнаруживают практически только там, где выявлены и наиболее ранние стоянки с орудиями труда — в Эфиопии, Кении, Танзании, Южной Африке, т. е. в долинах, расположенных с севера на юг почти вдоль всего великого восточноафриканского рифта. Из пяти видов грацильных австралопитеков — Australopithecus afarensis, Australopithecus africanus, Australopithecus garhi, Kenyanthropus platyops, Australopithecus bahrelghazali — только последний обнаружен за пределами очерченной территории, хотя и близко к ней (Республика Чад, район Коро-Торо).
Для всех разновидностей грацильных австралопитеков было свойственно прямохождение, хотя строение конечностей сохраняло признаки приспособленности к лазанию по деревьям и походку их, строго говоря, нельзя назвать подлинно человеческой. Прямыми свидетельствами прямохождения являются, в частности, данные археологического изучения цепочки застывших в вулканическом пепле следов ног на местонахождении Летоли в Танзании. Эти следы были оставлены 2,6 млн лет назад. Они сохранились до наших дней, благодаря тому что, после того как здесь прошел гоминид, дождь превратил слой пепла с отпечатавшимися на нем следами ног в прочный туф. Рост представителей описываемого рода гоминид достигал полутора метров. Зубная система была ближе к человеческой U-образной, в отличие от обезьяньей V-образной. По объему мозга (400–500 см3) австралопитеки практически не отличались от человекообразных обезьян. Максимальный размер черепной коробки, зафиксированный для австралопитековых гоминид, составляет 513 см3. Считается, что на ранних этапах истории гоминид прогресс в увеличении головного мозга был незначительным, если он вообще имел место. То, что на протяжении нескольких миллионов лет у австралопитеков не происходит заметных сдвигов в увеличении (а значит, и в развитии) головного мозга, с очевидностью свидетельствует о том, что они не обладали таким сугубо человеческим свойством, как систематическая трудовая деятельность. Следовательно, не приходится ожидать и открытия когда-либо в будущем археологических памятников со сколько-нибудь выразительным набором орудий, изготовленных австралопитеком.
Согласно палеоантропологическим обобщениям, которых придерживается большинство современных исследователей, в процессе развития человеческого рода на исторической арене последовательно выступали четыре главных действующих вида: Homo habilis, Homo erectus, Homo heidelbergensis и Homo sapiens.
Homo habilis — первый представитель собственно человеческой линии биологической эволюции. Принадлежащие ему костные остатки были обнаружены сначала на территории Танзании (Олдувайское ущелье, 1959 г.), затем в Кении (озеро Туркана) и Южной Африке (пещеры Стеркфонтейн и Сварткранс). Нетрудно заметить, что география находок совпадает с распределением находок грацильных австралопитеков. Время существования описываемого вида охватывает рамки примерно от 2,4 млн лет назад до примерно 1,7 млн лет назад. В течение указанного времени Homo habilis на занятой им территории в течение различных отрезков времени сосуществует по крайней мере с тремя видами австралопитеков.
В морфологическом отношении рассматриваемая группа гоминид обнаруживает внутри себя заметную таксономическую вариабельность. Из-за малочисленности конкретных данных эти расхождения не находят однозначного объяснения, которое бы формулировалось как хронологические отличия или, например, как внутривидовой половой диморфизм. У антропологов существуют разногласия по вопросу о том, признавать ли этот вид единственным представителем рода Homo для всего хронологического отрезка, занимаемого habilis, или же выделять для этого времени две или даже три разные родственные формы.
Те исследователи, которые склонны оценивать различия в хабилисах как очень существенные, уверенно говорят о необходимости выделения как минимум двух видов данной разновидности гоминид — Homo rudolfensis и Homo habilis. Сторонники этой точки зрения считают правильным вести главную линию эволюции скорее от более ранней и более продвинутой в развитии формы Homo rudolfensis, чем от собственно Homo habilis.
Разногласия по таксономическим оценкам различий в палеоантропологических материалах, относящихся к Homo habilis, более или менее примиряет позиция, согласно которой говорить о хабилисе можно в широком и узком смыслах. В узком смысле данный вид предстает таким, как он был описан для первой олдувайской находки. В широком же смысле в него в качестве отдельной формы входит более ранняя по возрасту находка на оз. Туркана.
Эта находка получила название Homo rudolfensis. Как бы то ни было, если судить по современным данным, то кроме Homo habilis в широком смысле или какой-то его конкретной формы не остается больше никого, кто мог бы претендовать на роль прямого предка следующего, более развитого вида раннего человека.
Анатомически Homo habilis резко отличается от австралопитеков, хотя рост его не слишком превышал размеры австралопитека и посткраниальный скелет во многом оставался сходным со скелетом предшественника, объем мозга при этом был значительно больше — 640 см3. Здесь четко фиксируется показатель направленной эволюции в сторону усложнения мозга через увеличение его общего объема — черта, которая у австралопитеков не отмечается. Изменился не только объем черепа, но и его строение. Отмечается развитие тех фрагментов черепной коробки, которые связаны с местом расположения речевого центра мозга. Округляется затылок, утончаются сами черепные кости. Зубы уменьшаются в размерах, как и вытянутость вперед лицевой части. Рука и кисть с противопоставленным большим пальцем приобретают почти современный вид.
Homo erectus занимает центральное место в решении проблемы ранних предков современного человека. Этот таксон объединяет все разновидности гоминид, фигурировавшие раньше под названиями питекантроп, синантроп, телантроп, атлантроп. Возможность обнаружения ископаемых остатков описываемого вида и более или менее верное с сегодняшней точки зрения определение его места в систематике гоминид были спрогнозированы в XIX в. в рамках идей эволюционизма. Первыми по времени обнаружения остатками эректусов стала знаменитая находка в 1890 г. Эженом Дюбуа питекантропа на Яве. В Африке аналогичные находки были связаны с открытием в 1949 г. в пещере Сварткранс в Южной Африке ископаемых костей телантропа капского. В конце 20-х годов XX в. в Китае были открыты и диагностированы выразительные остатки синантропа.
Последние и наиболее впечатляющие находки Homo erectus были сделаны в 80-х годах прошлого века в Восточной Африке. Самой известной из них является обнаружение полного скелета на стоянке Нариокотоме III на западном берегу оз. Туркана (Рудольф). Кости принадлежали мальчику в возрасте 12 лет. Датируется скелет возрастом 1,64 ± 0,03 млн лет назад. Рост мальчика должен был равняться 168 см; во взрослом состоянии — не менее 170 см. Рядом со скелетом не обнаружено ни одной археологической находки. По общему облику мальчик из Нариокотоме не очень сильно отличался от современного человека. Походка его была уверенно двуногой при полностью выпрямленном туловище. Пропорции тела, соотношения длины конечностей выглядели уже вполне человеческими. Тем не менее череп имел признаки, характерные именно для вида Homo erectus, — массивные надбровные дуги, покатый лоб, небольшой объем мозга (880 см3), отсутствие подбородочного выступа.
Очень ранние по датировкам (около 1,8 млн лет) остатки эректусов обнаружены и на восточном берегу оз. Туркана. По возрасту к ним близки находки из Эфиопии. В археологическом контексте с культурой олдувайского типа кости скелета Homo erectus найдены на стоянке Чесованья (Кения). На этом памятнике, который датируют временем около 1,4 млн лет назад, обнаружены, в частности, следы использования огня.
Одними из самых ранних (1,7–1,8 млн лет назад) являются находки Homo erectus и на стоянке Дманиси в Грузии. Они интересны как показатель распространения описываемого вида гоминид за пределы Африки и достаточно далеко в столь раннее время. Не менее важно то, что Homo erectus являлся здесь носителем культуры самого архаичного — олдувайского, а не ашельского типа. Высказывалось мнение о необходимости отнесения палеоантропологических находок из Дманиси к особой разновидности (Homo georgicus) с рассмотрением последнего как звена, промежуточного между габилисом и эректусом. На европейском континенте (Испания, Италия) наиболее ранние из известных находок эректусов относятся ко времени около 0,8 млн лет назад.
Хронологическая широта и территориальная распространенность Homo erectus не могли не привести к обсуждению проблемы таксономической однородности форм, объединяемых в этот вид. В результате была предпринята попытка разделить эректусов на два вида — африканский и азиатский. В ходе этих дискуссий в середине 70-х годов XX в. из общего ствола Homo erectus в качестве особого таксона был выделен Homo ergaster (в пер. с греч.: «человек работающий» — по смыслу эквивалент Homo habilis).
Сторонники теории замещения исходили из того, что Homo ergaster является общим предком как африканского, так и азиатского эректуса. Предполагалось, что второй впоследствии сформировался в особый вид. Такое разделение не всеми признавалось обоснованным. Сомнения в его правомерности усилились после обнаружения в Боури (Эфиопия) черепа Homo erectus возрастом 1 млн лет. Сходство морфологических признаков этой находки с черепами азиатского Homo erectus поставили под сомнение правомерность выделения Homo ergaster. С учетом этих и других данных большинство исследователей рассматривают обе названные разновидности в рамках единого таксона Homo erectus, который понимается в широком смысле вместе с Homo ergaster или в узком — без последнего. Это объясняется тем, что появление новых материалов на европейском и азиатском материках приводит часть палеоантропологов к заключению о представленности здесь не одной только, а обеих рассматриваемых разновидностей эректусов. Это, в частности, лишает основательности и высказывавшуюся идею об обратной миграции из Азии в Африку раннего человека в виде азиатского Homo erectus.
В качестве одного из ранних видов гоминид Европы рассматриваются палеоантропологические находки из Испании. В геологических отложениях в местности Гран Долина в Атапуерке, которые датируют временем около 800 тыс. лет назад, было найдено около сотни костей, относящихся к черепу, зубной системе, а также к посткраниальному скелету человека. Этим находкам был присвоен статус нового вида и дано название Homo antessesor («человек-предок»). Другие антропологи относят эти кости к эректусу, морфологически трудно отличимому от Homo ergaster. Существует мнение также о необходимости отнесения данных находок, равно как и палеоантропологических материалов из итальянской стоянки Чепрано, с индустрией доашельского облика к виду Homo heidelbergensis в начальной стадии его формирования.
Homo heidelbergensis как особый вид ископаемого человека был определен по находкам близ г. Гейдельберг в Германии в начале XX в. Специфика скелетных останков (особенно зубной системы этой древнейшей для Европы находки) заметно отличала их от уже известного к тому времени Homo erectus. Но говорить что-нибудь определеннее констатации большей продвинутости гейдельбергского человека в сторону современного человека в течение многих десятилетий оставалось затруднительным. В 30-х и 40-х годах XX в. остатки архаичных черепов различной сохранности были открыты в местностях Сванскомб в Англии, Штейнгейм в Германии и Фонтешвад во Франции. На всех них отмечалось такое же, как и на челюсти из Мауэра близ Гейдельберга, сочетание признаков питекантропа (как тогда называли эректусов) с современным человеком. Эта особенность явилась основанием для возникновения концепции пресапиенса.
Сторонники названной концепции обосновывали возможность существования прямой эволюционной линии от питекантропа к современному человеку через промежуточную форму пресапиенса, минуя неандертальцев. К пресапиенсам было отнесено несколько новых в то время европейских палеоантропологических находок, в том числе выразительные скелетные остатки из стоянок Кон де л’Араго во Франции, пещеры Петралона в Греции, Вертешселлош в Венгрии, Бильцигслебен в Германии, Кастель ди Гвидо и Скарио в Италии. Сюда же относили и одну кавказскую находку — фрагмент челюсти человека из пещеры Азых в Нагорном Карабахе.
Все пункты, с которыми связаны рассматриваемые палеоантропологические материалы, имеют более или менее единую археологическую характеристику как в стадиальном, так и в культурном отношениях. В преобладающем большинстве случаев эти памятники относятся к раннему или среднему ашелю с датировками в диапазоне примерно 700–400 тыс. лет назад. Однако известны и более поздние находки описываемого таксона, относящиеся к позднему ашелю и определенные как поздний вариант Homo heidelbergensis с датировками около 150 тыс. лет назад (грот Лазаре и навес Абри Сюар во Франции).
Таким образом, в эпоху раннего ашеля на европейском Средиземноморье и в Средней Европе, а возможно, и на Кавказе, был широко расселен вид раннего человека Homo heidelbergensis, сочетавший в себе морфологические черты эректуса и сапиенса. К чертам, сближающим его с современным человеком, относятся слабая выраженность скуловых костей лицевого отдела, высокий свод черепа, округлая форма затылка, отсутствие черепного гребня, заметный прогиб основания черепа, что связывается с выраженностью речевой функции.
Обобщая данные, касающиеся места Homo erectus и Homo heidelbergensis в филогенезе человека, можно указать на два устоявшихся подхода. Согласно теории полицентризма (мультирегионализма, многолинейного филогенеза), Homo ergaster расщепляется на два вида — Homo antessesor в Европе и Африке и Homo erectus в Азии. Homo antessesor, представленный гейдельбергским человеком в Европе и Homo rodesiens в Африке, эволюционировал соответственно в неандертальца (в Европе) и в сапиенса (в Африке). Что же касается Азии, то эректусы здесь доживают до времени появления неандертальцев в Европе и сапиенсов в Африке; в дальнейшем они эволюционируют в Homo daliensis (континентальный юго-восток Азии) и Homo soloensis (Меланезия).
Большинство ученых, однако, придерживается в настоящее время идеи однолинейной эволюции, или теории замещения. Homo ergaster (африканский Homo erectus) признается сторонниками этой концепции единым предком всех плейстоценовых гоминид. Позже на европейском и африканском континентах Homo ergaster/Homo erectus эволюционировал в Homo heidelbergensis. Европейская ветвь гейдельбержцев привела к Homo neanderthalensis, а африканская — к Homo sapiens. В дальнейшем сапиенс заместил собой в Европе неандертальцев, а в Азии — гейдельбержцев и/или поздних эректусов.
Homo sapiens — современный вид человека — результат предшествовавшей эволюции рода Homo, которая длилась не менее двух миллионов лет. Согласно данным генетики, все современное человечество ведет начало от одной небольшой популяции, которая сформировалась на африканском континенте около 200 тыс. лет назад. Из приведенных выше данных видно, что эта предковая общность сложилась в недрах предшествующего вида Homo erectus. Спустя 50-100 тыс. лет после своего появления новый вид анатомически современного человека населяет новые обширные территории за пределами африканского континента. Примерно 40–50 тыс. лет назад сапиенс заселяет практически весь Старый свет.
Преобладающее большинство современных палеоантропологов полагает, что биологическая история Homo sapiens началась на африканском континенте примерно 250 тыс. лет назад. Иногда выделяют два этапа в становлении данного вида — стадии раннего и позднего архаичного сапиенса. К ранним относятся материалы известных местонахождений Омо 2 в Эфиопии с возрастом около 130 тыс. лет назад, Элие-Спрингс (дата в диапазоне от 100 до 200 тыс. лет назад) на оз. Туркана (Кения), Флорисбад в Южной Африке (возраст — 259 тыс. лет назад) и др. Все эти находки по времени следуют за самыми поздними остатками африканской ветви представителей Homo heidelbergensis. Сохраняя тесное морфологическое сходство с последними, они обнаруживают в то же время качественный прогресс в направлении к сапиенсу. Это дает основания для вывода о прямой филогенетической линии, ведущей от тропической разновидности гейдельбергского человека к сапиенсу.
Развитая стадия архаичного сапиенса лучше всего иллюстрируется палеоантропологическими материалами, происходящими с Ближнего Востока. Наиболее известными в этом отношении являются скелетные остатки из пещер Схул и Кафзех. В отложениях с мустьерской культурой в пещере Мугаррет эс-Схул (гора Кармел в Израиле) были обнаружены костные остатки десяти индивидов разного возраста и пола. Датировки слоев с указанными находками противоречивые. Сейчас их возраст оценивается ориентировочно в 100-80 тыс. лет назад. Если судить по наиболее репрезентативному скелету, то человек из пещеры Схул предстает высоким, с пропорциями тела современного человека, большим объемом мозга (1518 см3), низким широким лицом с небольшой высотой орбит, выраженным подбородочным выступом и округлым затылком. При этом сохраняются архаические признаки в виде заметно выступающих надбровных дуг и наклоненного назад лба.
Как показывают археологические данные, сапиенсы Ближнего Востока сосуществовали во времени с представителями вида Homo neanderthalensis. Многочисленные костные остатки последних выявлены вместе с выразительными археологическими материалами в слоях пещер Кебара, Амуд (Израиль), Шанидар (Иракский Курдистан). Наличие неандертальских признаков у сапиенсов Леванта иногда объясняют возможностью метисации двух указанных видов. Предполагают, что одним из следствий этого могло стать дальнейшее ускорение на обширной территории выработки всего набора морфологических признаков, свойственных для современного Homo sapiens.
Вопрос о том, когда и как сформировавшийся Homo sapiens освоил различные регионы Европы и Азии, остается до конца не выясненным. Очевидно, что неандертальцы обитали на обоих континентах примерно до 30–35 тыс. лет. На территории современной Западной Европы к этому времени на местном субстрате начинает складываться культура, характерная для новой, следующей за мустьерской, культуры (верхний палеолит). Создавала эту культуру (культура Шательперрон, Франция) популяция местной, европейской разновидности неандертальцев. Но несколько раньше, около 40 тыс. лет назад, в Южной Европе появляется новая — ориньякская культура, которая по своему облику уже не является мустьерской. Вскоре она распространяется от Атлантики до Урала и даже шире, включая часть Северной Евразии.
В вопросе об ориньякской культуре в данном случае наиболее интересно то, что ее носителем определенно был Homo sapiens. Следовательно, если на всем европейском континенте культура эпохи мустье в определенный момент довольно стремительно сменяется верхнепалеолитической — ориньякской, то возникает вопрос о том, не сменился ли и антропологический тип самого человека? Конкретные палеоантропологические материалы отвечают на этот вопрос положительно. Смена мустьерской культуры верхнепалеолитической в антропологическом свете предстает как вытеснение вида Homo neanderthalensis видом Homo sapiens. Процесс этот не был кратким, одномоментным. Популяции неандертальцев на некоторых территориях (например, Крым, Кавказ) существовали еще по крайней мере 10 тыс. лет, после того как первые ориньякцы появились в Европе.
Приведенное выше описание относится к сценарию, называемому теорией моноцентризма, или теорией замещения. Как говорилось выше, этого взгляда придерживается большинство современных палеоантропологов и археологов. Но существует и другой вариант объяснения данного процесса. Согласно второй теории — мультирегионализма — процесс накопления у ранних гоминид анатомически современных черт (сапиетизация) в разных регионах протекал самостоятельно. В этой связи одной из самых дискуссионных остается проблема места Homo soloensis в систематике поздних представителей человеческого рода. Отношение к этой проблеме очерчивает линию разделения между сторонниками теорий моноцентризма и полицентризма.
Первые находки Homo soloensis были сделаны в 1930 г. на Яве в местности Нгандонг. В отложениях одной из террас реки Соло среди тысяч фаунистических остатков оказалось 12 фрагментов скелета человека. Индивид, которому принадлежали эти кости, обладал объемом мозга гораздо большим, чем яванский питекантроп. Соотношение это составляло 1200 к 883 см3. Однако по другим существенным морфологическим признакам (например, массивные надбровные дуги, значительная толщина костей черепа) человек из Соло оказывался ближе к Homo erectus. Принимая во внимание это, а также учитывая время его существования (200-25 тыс. лет назад), Homo soloensis рассматривается как хронологический эквивалент европейских неандертальцев. По существу же, ему отводится место представителя одной из позднейших популяций и яванского питекантропа. Он синхронен с Homo sapiens, но отличен от него в видовом отношении.

Направление первоначального расселения человека на территориях Африки и Азии
На основании сравнения находок из Нгандонга с палеоантропологическими материалами с озера Вилландра в Австралии (датировка — 15 тыс. лет назад) сторонники полицентрической теории считают Homo soloensis промежуточным звеном, связывающим позднейшего Homo erectus с анатомически современным человеком австралийского аборигенного типа.
Для Индокитая (находки на континентальном юго-востоке Китая) вопрос о появлении анатомически современного вида человека «мультирегионалисты» решают по той же модели, которая описана выше для Меланезии и Австралии. Место Homo soloensis здесь занимает другая постэректоидная форма, определяемая как вид Homo daliensis.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Антропогенез славян
Антропогенез славян В дальнейшем славяне, укрывшиеся в лесах Восточной и Центральной Европы, пострадали не столько от гуннов, аваров и прочих кочевников, сколько от финно-угров. Местное население было враждебно настроено по отношению к переселенцам из степей. Ариев,
Антропогенез: эволюция человека
Антропогенез: эволюция человека Антропогенез – происхождение и дальнейшее развитие человека разумного – является одной из острых и, пожалуй, при большом количестве работ на эту тему, одной из самых малоисследованных тем.Лишь в конце XIX столетия утвердилась концепция
Антонио Эспада, Сандомир Хлодвиг АНТРОПОГЕНЕЗ С ТОЧКИ ЗРЕНИЯ ТЕОРИИ ПРОСТРАНСТВЕННЫХ АНОМАЛИЙ
Антонио Эспада, Сандомир Хлодвиг АНТРОПОГЕНЕЗ С ТОЧКИ ЗРЕНИЯ ТЕОРИИ ПРОСТРАНСТВЕННЫХ АНОМАЛИЙ Вообще говоря, теория пространственных аномалий в основе своей сближается с материализмом. Главная ее особенность состоит в том, что ТПА считает материю и энергию не